Bot. Bull. Acad. Sin. (2001) 42: 39-44
Chiang et al. Fungal rDNA sequences from Miscanthus
Detection of leaf-associated fungi based on PCR and nucleotide sequence of the ribosomal internal transcribed spacer (ITS) in Miscanthus
Yu-Chung Chiang1,2, Chang-Hung Chou1, Pei-Ray Lee1, and Tzen-Yuh Chiang3,*
1Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan
2Department of Biology, National Taiwan Normal University, Taipei 100, Taiwan
3Department of Biology, Cheng-Kung University, Tainan 701, Taiwan
(Received November 23, 1999; Accepted July 10, 2000)
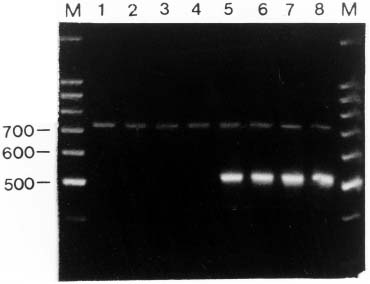
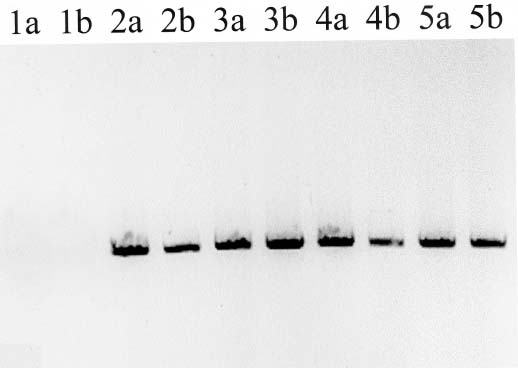
Abstract. PCR amplification of the internal transcribed spacer region (ITS) of the nuclear ribosomal DNA from the genomic DNA isolated from the leaf tissue of Miscanthus sinensis varieties and M. floridulus resulted in two amplified fragments. Via FASTA search, the shorter length of about 500 base pairs was identified as the ITS region of fungi symbiotic in the grass foliage. Two Cladosporium species, one Fusarium and one basidiomycete were added to the mycoflora of Miscanthus grassland. Using a Southern blotting and hybridization, fungal ITS was also detected from the genomic DNAs isolated from sterilized seedling, a suggestive seed transmission of endophytic fungi. The "contaminated" genomic DNA may be a hindrance to the phylogenetic analysis of Miscanthus itself, but may provide other insights into the biodiversity of grassland ecosystems. The endophytic foliage fungi were not detected in M. sinensis var. condensatus, probably a result of its apomictic reproduction.
Keywords: Contamination; Foliage fungi; Miscanthus sinensis; Mycoflora; Phylogeny; rDNA ITS.
Introduction
The association between fungi (both endophytic and epiphytic) and higher plants is thought to be ubiquitous (Clay, 1988; Legault et al., 1989; Petrini et al., 1992). Nevertheless, the prevalence of symptomless fungi on healthy plants has become clear only lately (cf. Camacho et al., 1997). Bills (1996) even suggested that endophytic fungi are basic components of microbiota and are distinct from pathogens ecologically. In grasslands, where Poaceae are dominant, symbiotic fungi, both endophytic and epiphytic, may play critical roles in plant-microbe interactions. Diverse taxa of fungi have been isolated and identified from the leaf tissue of many grasses (Harada, 1987; Nakase et al., 1987; Watson and Dallwitz, 1992).
Molecular techniques have been applied widely in ecological studies (e.g., Wirgin et al., 1997) and wildlife conservation (Avise and Hamrick, 1996; O'Corry-Crowe et al., 1997) in the past several years. In many studies the polymerase chain reaction (PCR) has proved powerful in detecting cryptic genetic variation, e.g., the DNA of symbionts that can not be cultured and separated from their cosymbionts (Haddad et al., 1995), and the microbial floristic composition (cf. Hansen and Hanson, 1996). For systematic and ecological studies, many universal prim
ers have been designed, which may be used across a large range of taxa (Dumolin-Lapegue et al., 1997; Chiang et al., 1998; Chow and Hazama, 1998). The primer sequences of the internal transcribed spacer (ITS) region of the nuclear ribosomal DNA have been widely used for resolving phylogenetic relationships at the species or generic levels (White et al., 1990; Baldwin, 1992). Two recent reports documented the amplification of fungal sequences from plant foliage (Liston et al., 1996; Klein and Smith, 1996). Employing phylogenetic analysis, Camacho et al. (1997) was able to identify the endophytic fungi in Picea based on the ITS nucleotide sequences. In addition, sequences of the nuclear ribosomal internal transcribed spacer region are known providing species-specific genotypes in many fungi (Egger, 1996; Norman and Egger, 1996; Holst-Jensen et al., 1997; Zhang et al., 1997; Vralstad et al., 1998).
A species complex of Miscanthus sinensis Anders. consists of several intraspecific taxa that occur in different elevations on Taiwan (Chou et al., 1999a). The history of adaptive radiation and secondary contacts in the species complex has been reconstructed based on genetic variation of both nrITS and cpDNA atpB-rbcL spacer sequences (Chou et al., 1999b). Most taxa play a dominant role in the floristic composition of grasslands at different elevations. Examples include M. sinensis var. transmorrisonensis (Hayata) Lee in high mountains, M. sinensis var. formosanus Hack. at middle elevations, and M. sinensis var. glaber (Nakai) Lee in lowlands. In contrast to these taxa with a wide range in distribution, M.
*Corresponding author. Tel: +886-6-2757575 ext. 65525; Fax: +886-6-2742583. E-mail: tychiang@mail.ncku.edu.tw