Bot. Bull. Acad. Sin. (2001) 42: 45-52
Lin et al. Host cell death induced by the c2 mutant of f1
Mechanism of host cell death induced by infection of Escherichia coli with the c2 clear-plaque mutant of phage f1
Shan-Hua Lin1, Wen-Ping Chen2, and Tsong-Teh Kuo1,2,*
1Institute of Molecular Biology, Academia Sinica, Nankang, Taipei 11529, Taiwan, Republic of China
2Department of Botany, National Taiwan University, Taipei, Taiwan, Republic of China
(Received September 17, 1999; Accepted August 7, 2000)
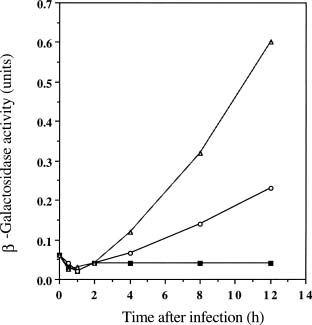
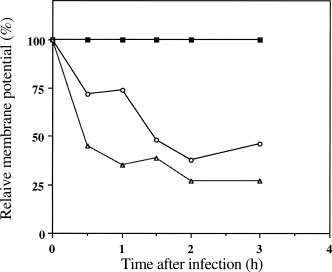
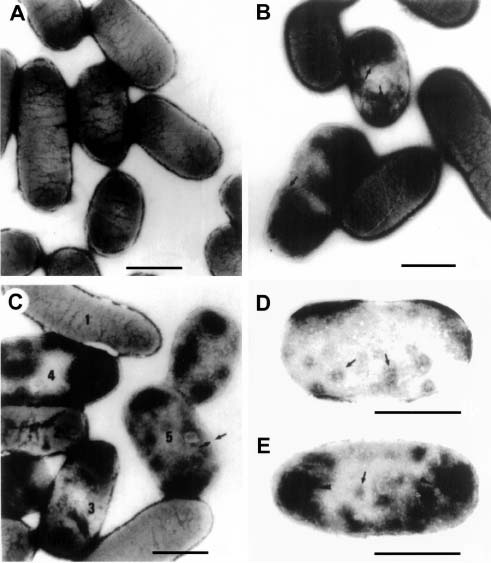
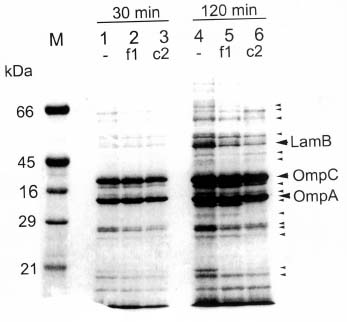
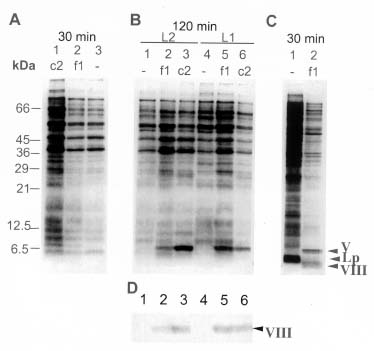
Abstract. The c2 clear-plaque mutant arose spontaneously from the turbid plaque-inducing wild-type strain of bacteriophage f1. The mechanism of host cell death induced by infection of Escherichia coli with c2 has now been investigated. A marked decrease in cell membrane potential was apparent as early as 30 min after infection with c2, and leakage of cell contents was apparent after 4 h. Transmission electron microscopy also revealed the accumulation of granular membrane-like structures within cells at early stages of c2 infection. Electrophoretic analysis showed that the abundance of several bacterial outer membrane proteins was markedly reduced 2 h after infection with c2. Furthermore, substantial amounts of the phage coat protein (gpVIII) and single-stranded DNA-binding protein (gpV) were apparent in the inner membrane of c2-infected cells 2 h after infection. These data support the hypothesis that the death of c2-infected cells results from phage-induced damage to the bacterial cell membrane.
Keywords: Cell death; Clear plaque; Escherichia coli; Filamentous phage f1.
Introduction
Escherichia coli infected with the bacteriophage f1 continues to grow and divide, although at a slower rate than that of uninfected bacteria (Marvin and Hohn, 1969). In contrast, infection of nonpermissive bacteria with f1 mutants containing amber mutations in certain of the eight phage genes results in a marked decrease in cellular DNA, RNA, and protein synthesis (Hohn et al., 1971a,b) and in the cessation of cell division (Pratt et al., 1966). Furthermore, such mutants increase cell membrane permeability as well as induce leakage of cell contents and cell death at early stages of infection (Hohn et al., 1971a,b). Electron microscopy has revealed that absorptive infection of E. coli with f1 mutants containing amber mutations in genes I, III, IV, V, or VI results in the accumulation of multiple folded membranous structures within the bacteria, especially at the two poles of the cell (Schwartz and Zinder, 1967; Ohnighi, 1971). These changes induced in bacteria by f1 mutants result from the accumulation of large amounts of the gene VIII-encoded protein. Thus, infection of host cells with gene VIII amber mutants does not induce such changes in the structure of cell membranes (Woolford et al., 1974).
We previously isolated a spontaneous mutant, c2, of wild-type f1 that exhibits a clear-plaque phenotype (Kuo et al., 2000). Infection with this mutant results in the synthesis of increased amounts of both phage DNA and gpII compared with those apparent in cells infected with wild-
type f1 (Chen, 1980). We have now investigated the time courses of the various changes in bacterial cell morphology and physiology induced by infection with c2 in order to shed light on the mechanism of host cell death.
Materials and Methods
Preparation of Phage-Infected Bacterial Cultures
Escherichia coli K38 and wild-type phage f1 were obtained from Dr. N. D. Zinder of Rockefeller University. The clear-plaque mutant c2 arose spontaneously from wild-type f1. The bacteria were grown to log phase at 37°C in LB medium with shaking. Phage were harvested from the supernatant of bacterial cultures that had been incubated for ~6 to 10 h after infection at a phage multiplicity of 10.
For the assay of b-galactosidase activity, the bacteria were grown in LB medium containing 1 mM isopropyl-b-D-thiogalactopyranoside. Bacterial cultures at a density of ~1 × 108 to 5 × 108 cells/ml were infected with phage at a multiplicity of 10. When indicated, chloramphenicol was added to the phage-infected cultures at a final concentration of 30 µg/ml at 0, 40, 90, or 360 min after infection.
Assay of b-Galactosidase Activity
The activity of b-galactosidase was assayed as described by Miller, 1971. The amounts of cellular and extracellular enzyme activity were determined by first separating bacterial cultures into a bacterial pellet and the culture supernatant by centrifugation at 10,000 g for 5 min. Portions (1 ml) of the supernatant and resuspended bacterial pellet were incubated at 20°C for 5 min prior to the addition of 50 µl of o-nitrophenyl-b-D-galactoside (4 mg/
*Corresponding author. Tel: +886-2-27899213; Fax: +886-2-27826085; E-mail: tehkuo@sinica.edu.tw