Bot. Bull. Acad. Sin. (2001) 42: 53-60
Chang et al. Warm roots and cold acclimation provide protection from chilling injury
Cold-acclimation and root temperature protection from chilling injury in chilling-sensitive mungbean (Vigna radiata L.) seedlings
Mon-Yaw Chang, Swu-Ling Chen, Chung-Fen Lee, and Yih-Ming Chen*
Department of Botany, National Taiwan University, Taipei 106, Taiwan, Republic of China
(Received September 8, 1999; Accepted June 21, 2000)
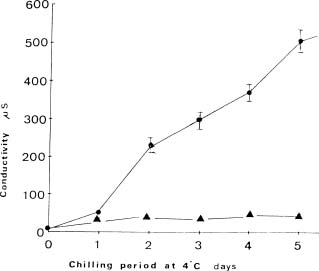
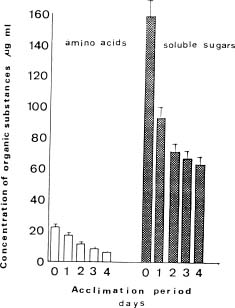
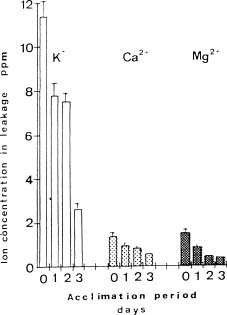
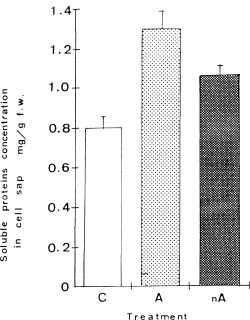
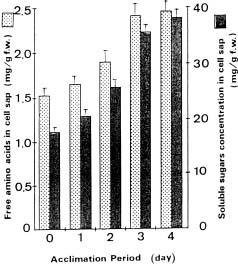
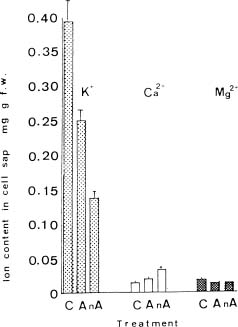
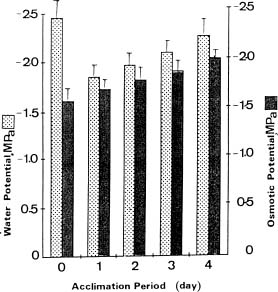
Abstract. Exposure of mungbean seedlings to 4°C for 2 days induced irreversible chilling injury. The major cation in the leakage from tissues of unacclimated seedlings was K+, the loss of which was 7 to 10-fold greater than that of Ca++ or Mg++. Acclimation of seedlings at 10°C protected them from the injuries caused by the 4°C treatment. Acclimation of seedlings at 10°C for 2 to 3 days, significantly decreased the conductivity and the concentration of soluble sugars, free amino acids, and cations (K+, Mg++ and Ca++) in the leakage. Compared to the 28°C-root/28°C-shoot control seedlings, those in the 28°C -root/4°C -shoot treatment did not suffer noticeable injury, but seedlings in the 4°C-root/4°C-shoot treatment did. The solute potential, water potential, and the concentration of free amino acids and cations (K+, Mg++, and Ca++) in the cell sap of the 28°C-root/4°C-shoot seedlings were similar to those of the control seedlings.
Keywords: Cell sap; Chilling injury; Cold acclimation; Conductivity; Leakages; Mungbean (Vigna Radiata L.); Root temperature.
Introduction
Tropical and subtropical plants exhibit marked physiological and biochemical dysfunctions, commonly referred to as chilling injury, when they are exposed to temperatures below l0°C to l2°C (Graham and Patterson, 1982; Wang, 1982; Guy, 1990). These dysfunctions include alteration in membrane structure and lipid composition (Lyons and Raison, 1970), metabolic modifications (Sochanowicz and Kaniuga, 1979; Levitt, 1980; Trevanion et al., 1995), changes in protein content (Marmiroli et al., 1986; Bredenkamp and Baker, 1994) and enzyme activities (Byrd et al., 1995; Kumar and Triphathy, 1998), phosphorylation of thylakoid proteins (Bannett, 1991), cyclosis (Lewis, 1956), redistribution of intracellular calcium ions (Bush, 1995), cellular leakage of electrolytes and amino acids, and a diversion of electron flow to alternate pathways (Leopold and Musgrave, 1979). Dysfunctions associated with chilling stress in mungbean seedlings may be attributable to the alteration of gene expression (Guy et al., 1985; Mohapatra et al., 1989; Kurkela and Franck, 1990; Ouellet et al., 1993; Wolfraim and Dhindsa, 1993; Wolfraim et al., 1993; Hughes and Dunn, 1996; Kung et al., 1998). However, there is still a paucity of information on the effects of cold acclimation and root temperature on chilling injury. In this study, we provided protection from chilling injury in mungbean seedlings using cold acclimation and warm root treatments.
Materials and Methods
Plant Material
Mungbean (Vigna radiata L.) seeds were immersed in running water overnight and then planted in vermiculite in a growth chamber set at 28°C and 45 µmol m-2 sec-1 (14L/10D). The seedlings were grown for 5 days under these conditions and then separated into two groups. Seedlings in the chilling treatment group were transferred to 4°C and 45 µmol m-2 sec-1 for various lengths of time. The control group seedlings were maintained at 28°C and 45 µmol m-2 sec-1. For the cold-acclimation process, 5-day-old seedlings were subjected to l0°C for various lengths of time and then transferred to 4°C. To study the effect of root temperature on chilling injury, seedlings were grown at either 28°C or 4°C in a water culture containing Hoagland solution (Epstein, 1972).
Procedures
Preparation of cell sap from leaf discs and measurement of conductivity. Cell sap was obtained from frozen and thawed leaf discs as described by Zudo et al. (1983). The conductivity of electrolyte leakage in the cell sap from leaf discs was used as a measure parameter of chilling injury (Sukumaran and Weiser, 1972).
Quantitative analysis of soluble sugars, amino acids, proteins and cations. The concentration of amino acids that leaked from leaf discs into the incubation medium was determined by the ninhydrin methods described by Moore and Stein (1954), using leucine as a standard. Soluble sugars in the incubation medium were measured by the
*Correspondence author. Tel: 02-23630231 ext. 2360; Fax: 02-23919096; E-mail: yihmingc@ccms.ntu.edu.tw