Bot. Bull. Acad. Sin. (2001) 42: 61-66
Liu et al. Karyomorphology of Biebersteinia Stephan
Karyomorphology of Biebersteinia Stephan (Geraniaceae) and its systematic and taxonomic significance
Jianquan Liu1,2,*, Tingnong Ho1, Shilong Chen1, and Anmin Lu2
1Northwest Plateau Institute of Biology, the Chinese Academy of Sciences, Xining 810001, China
2Plant Systematic and Evolutionary Laboratory, Institute of Botany, the Chinese Academy of Sciences, Xiangshan, Beijing 100093, China
(Received January 21, 2000; Accepted May 29, 2000)
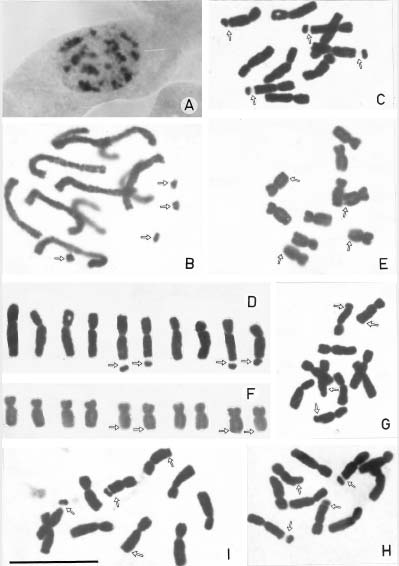
Abstract. The systematic and taxonomic position of Biebersteinia Stephan has long been in dispute. The present paper describes for the first time the karyomorphology of two species in Biebersteinia Stephan. Both species commonly showed the interphase nuclei of the simple chromocenter type and the mitotic prophase chromosomes of the interstitial type. The karyotype formulae of both B. heterostemon and B. odora were 2n=10=2m(2sec)+8sm(2sec), belonging to the 3A type of Stebbins' classification. The karyotype of this genus is recorded for the first time. The basic chromosome numbers of four of the five known species of Biebersteinia have been recorded as x=5. The combination of resting nuclei of the simple chromocenter type, mitotic prophase chromosomes of the interstitial type, two pairs of chromosomes with four obvious secondary constrictions at the mitotic prophase and metaphase stages, and the peculiar 3A karyotype in Biebersteinia can be regarded as the karymorphological marker of this genus. The karyomorphological data presented here do not support the traditional grouping of this genus in Geraniaceae. The unique karyomorphology of Biebersteinia justifies its familiar or ordinal status, which is congruent with embryological, anatomical, chemical and molecular data. The systematic position of Biebersteinia needs further study.
Keywords: Biebersteinia; Karyomorphology; Systematics.
Introduction
Biebersteinia Stephan, comprising five species: B. heterostemon Maxim, B. odora Stephan ex Fisch, B. multifida DC., B. orphanidis Boiss. and B. leiosepala, is distributed in mountainous, semi-arid areas of central and western-Asia to the eastern Mediterranean (Knuth, 1912; Yannistaros et al., 1996). The systematic and taxonomic position of Biebersteinia Stephan has long been in dispute. Stephan (1806) positioned Biebersteinia between Grielum L. (Neuradaceae) and Suriana L. (Simaroubaceae) when establishing the genus. Later, De Candolle (1924) suggested a position of Biebersteinia in `Zygophylleae spuriae alternifoliae' and possible affiliations with Rosaceae, Geraniaceae and Rutaceae. It was Endlicher (1841) who raised Biebersteinia to a family status for the first time. But Hooker (1875) treated it as a tribe in Potentillae in the Rosaceae. However, Boissier (1867) included it as a genus of Geraniaceae. In the recent angiosperm systems, Thorne (1992) and Cronquist (1981, 1988) followed Boissier (1867) and Knuth (1912) and treated it as a genus of Geraniaceae while Dahlgren (1989) and Takhtajan (1987) recognized it as a family close to Geraniaceae. Takhtajan (1997) further raised it to an order near Geraniales. Most recently, Bakker et al. (1998) qualified its family status and implicated strong affiliation with
Sapindales based on the molecular data. This result was positively accepted by APG (1998) in their new angiosperm ordinal system.
In addition to the gross-morphological and molecular evidence, other data such as the pollen morpholgy, embryology, anatomy, chemistry and karyomorphology can also be of great importance in clarifying the systematic position of a taxon (Takhtajan, 1997). Bortenschlager (1967) studied the pollen morphology of Biebersteinia, and suggested the genus should be recognized as a family near the Potentillae of Rosaceae. Nevertheless, the phanerothetic discoid nectaries of Biebersteinia were very similar to those found in most Geraniaceae (Link, 1990). But the presence of anacampylotropous ovules (Kamelina and Konnova, 1990), the lack of a persistent elongated central column in the ovary and a reduced tegmic seed coat (Boesewinkel, 1997), and other chemical data (Bate-Smith, 1973) distinguish Biebersteinia from all taxa assumed to be related to it.
In spite of the accumulating molecular data, chromosome information continues to be important in assessing phylogenetic relationships (Carr et al., 1999). Concerning the karyomorphological data of Biebersteinia, only the chromosome numbers of B. multifida DC. (Aryavand, 1975) and B. orphanidis Boiss. (Constantinidis, 1996) were reported to be x=5. The karyotype, the interphase nuclei, and the mitotic prophase chromosomes have never been reported. In the present paper, the chromosome numbers
*Corresponding author. E-mail: ljqdxy@public.xn.qh.cn