Bot. Bull. Acad. Sin. (2001) 42: 67-84
D'Arcy et al. The genus Tubocapsicum (Solanaceae)
The genus Tubocapsicum (Solanaceae)
William G. D'Arcy1, Richard C. Keating1, Zhi-Yun Zhang2, and Ching-I Peng3,*
1Missouri Botanical Garden, P.O. Box 299, St. Louis, MO 63166, USA
(William G. D'Arcy passed away on December 16, 1999)
2 Institute of Botany, Chinese Academy of Sciences, Xiangshan, Nanxincun 20, Beijing 100093, China
3Institute of Botany, Academia Sinica, Taipei 115, Taiwan
(Received November 23, 1999; Accepted June 22, 2000)
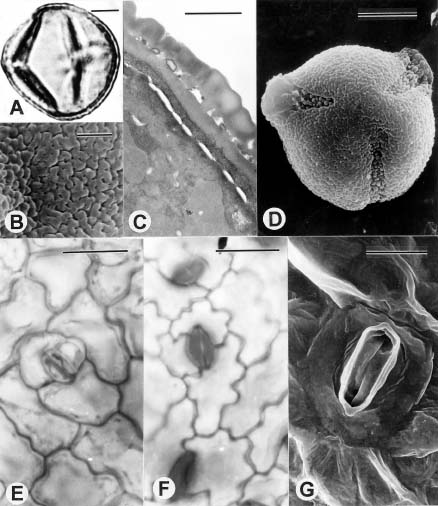
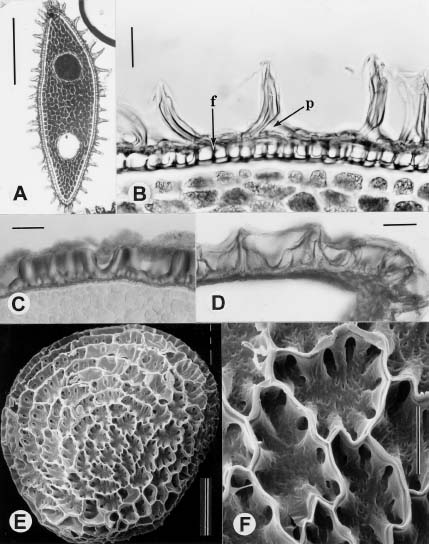
Abstract. Tubocapsicum (Solanaceae: Solanoideae), a genus of two Asian species, was formerly included in Capsicum. Tubocapsicum anomalum is a herb with small yellow flowers and red berries that is found in Japan, southern China, Taiwan, and the Philippines; T. obtusum is found in Japan. A general taxonomic description is complemented by new detailed descriptions and illustrations of growth morphology, vegetative histology and stomata, floral structure, vasculature, fruit structure, seeds, and pollen. On the basis of this analysis as well as cpDNA investigations by of Olmstead et al. (1999), the genus Tubocapsicum appears to be most closely related to subfamily Solanoideae genera Aureliana and Withania.
Keywords: Anatomy; Anther; Branching; Capsicum; Floral vasculature; Fruit; Growth pattern; Nectar; Pollen; Revision; Seeds; Solanaceae; Stomata; Taxonomy; Tubocapsicum.
Introduction
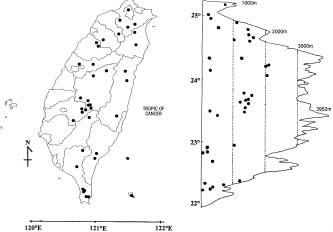
The genus Tubocapsicum Makino is confined to eastern Asia (Figures 1, 2). It embraces one widespread species, T. anomalum (Franch & Sav.) Makino and a poorly known second species, T. obtusum (Makino) Kitamura, of Japan. The genus is not well known since there has been little published information, and there are few specimens in western herbaria. In order to redress this obscurity, we undertook a study of living plants of the genus cultivated from seeds from Taiwan, as well as herbarium material from throughout its range. Here we report our observations and provide a treatment of the genus Tubocapsicum with descriptions, drawings, photos, and an analysis of its possible relationships within the Solanaceae. Nearly all of the observations on the structure and biology of the genus to follow are of T. anomalum as T. obtusum is only known thus far from a few herbarium specimens.
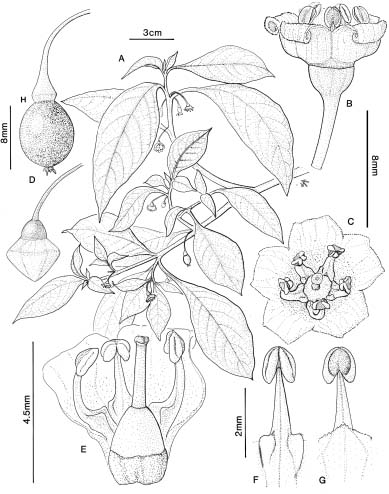
Plants of Tubocapsicum anomalum (Figures 3, 4, 5, 6) are erect or sprawling herbs with entire leaves, few-flowered inflorescences, small campanulate flowers, and red juicy berries with discoid seeds. In most features, they resemble members of Acnistus Schott, Aureliana Sendt., Capsicum L., Vassobia Rusby, Witheringia L'Her. and other genera of the American tribe Capsiceae. However, as discussed below, seed comparisons and cpDNA studies (Olmstead et al., 1999) argue for separating Tubocapscium and Aureliana from tribe Capsiceae, along
with Withania Pauq. which has been placed in tribe Physaleae.
In this paper we present a summary of our observations of living plants and herbarium material of Tubocapsicum anomalum. We include a report on germination and growth patterns, vegetative histology and vasculature, floral structure and vasculature, fruit structure, pollen, seeds, quality of the nectar, notes on its cultivation, and a general discussion of how it differs from other genera. We conclude with a systematic revision based on literature and herbarium collections including nomenclature of the genus, its two species, and a map of its distribution.
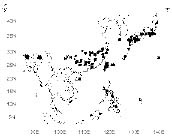
Figure 1. Distribution of Tubocapsicum anomalum. Solid squares indicate specimens seen by the authors. Large circles indicate reports from the literature.
*Corresponding author. E-mail: bopeng@gate.sinica.edu.tw