Bot. Bull. Acad. Sin. (2001) 42: 93-100
Lai et al. Genetic relationships in tea
An assessment of genetic relationships in cultivated tea clones and native wild tea in Taiwan using RAPD and ISSR markers
Jou-Ann Lai, Wei-Chen Yang, and Ju-Ying Hsiao*
Department of Botany, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received May 2, 2000; Accepted October 27, 2000)
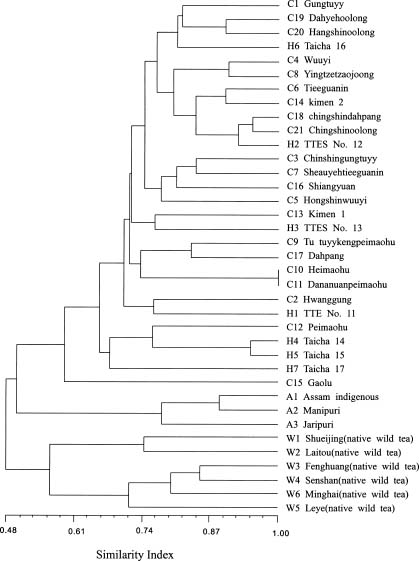
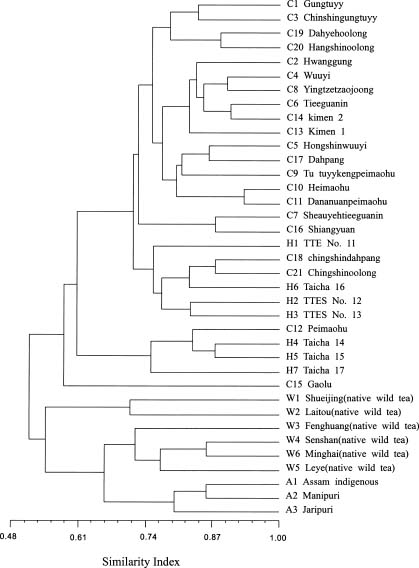
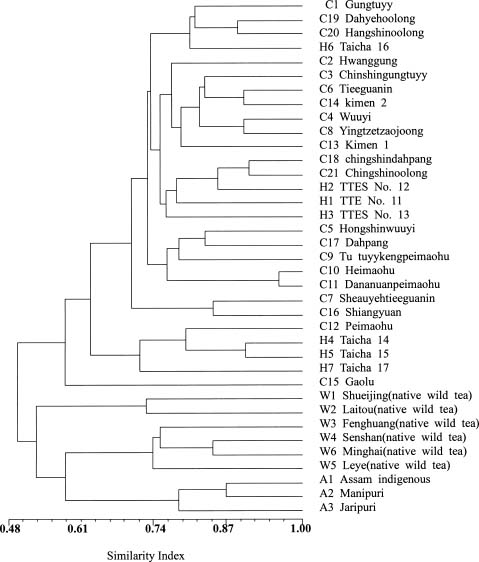
Abstract. Tea (Camellia sinensis [L.] O. Kuntze) is an important beverage crop in Taiwan. Most of the cultivated clones were introduced from China and India though some native wild teas are distributed in the mountains of central and southern Taiwan. In this study, 37 tea samples were evaluated using RAPD and ISSR markers. The samples comprised 21 clones of China, 3 clones of Assam, 7 hybrid clones between China and Assam tea, and 6 individual samples of native Taiwanese wild tea. A total of 53 and 56 polymorphic RAPD and ISSR markers respectively, were scored. The results of cluster analysis based on RAPDs revealed that three major groups could be recognized, i.e., cultivars of China tea and the cultivars developed in Taiwan from hybridization and selection; Assam tea; native Taiwanese wild tea. The native Taiwanese wild teas were, however, most distant in the clustering tree. In the ISSR dendrogram, Taiwanese native wild teas clustered closely with Assam tea then with China tea and the Taiwanese hybrid cultivars. The population gene diversity of the native wild tea was found to be the highest among the three populations studied. Analysis of molecular variance (AMOVA) revealed that the variance component within groups was larger than that among groups. The correlation coefficient between similarity matrices based on RAPD and ISSR was 0.811. A Mantel test revealed that the correlation was highly significant (p<0.001), indicating good congruence between the results of these two molecular markers.
Keywords: AMOVA; Camellia sinensis; Genetic variation; ISSR; RAPD; Tea.
Introduction
Tea (Camellia sinensis [L.] O. Kuntze) is an important beverage crop in Taiwan, having been cultivated on the island for more than two hundred years. The tea trade dates back to 1865. The export of tea from Taiwan has, however, been decreasing gradually since 1980 due to competition from other tea-exporting countries (Zuan and Shi, 1995). The production of high quality tea with regional characteristics has nevertheless remained a highly profitable business. The breeding of the new high quality varieties and the conservation of tea germplasm resources continue to be important for the sustainable cultivation of tea in Taiwan.
Tea is an out-crossing species, and selected elite genotypes are propagated vegetatively and released as clonal varieties. Clonal identification has traditionally been based on morphological descriptors such as plant shape, leaf shape, young leaf type, and fruit shape. However, as in many out-crossing crops, tea is highly heterozygous with most of its morphological, physiological and biochemical descriptors showing continuous variation and high plasticity. Similarly, most morphological traits are influenced by environmental factors, plant age and phenology. Furthermore, since objectivity is crucial to accurate morphological typing, the above factors render the use of
such descriptors in plant identification and discernment of genetic relationships difficult. Molecular markers such as RAPD and ISSR can, however, be used for these purposes. The advantages of molecular markers are that they are least affected by environmental factors and are almost unlimited in number. They also offer a possibility to observe the genome directly, and thus eliminate the shortcomings inherent in a phenotype observation.
The first tea cultivar cultivated in Taiwan was introduced from Fukien (China), between 1796 and 1820. More cultivars were introduced from China thereafter. From 1926, some of the new cultivars were introduced from India. The cultivars introduced from China belong to China tea (var. sinensis [L] O. Kuntze) while those introduced from India belong to Assam tea (var. assamica [Mast.] Kitam.). The majority of tea clones cultivated in Taiwan are China tea, reflecting the order of introduction and the preference for domestic consumption. Besides the introduced clones, new clones have been developed by hybridization and selection. The native wild tea (var. assamica [Mast.] Kitam.f. formosensis Kitam.) grows in the central and southern mountains of Taiwan. Based on leaf characteristics and tree type, they have been taxonomically classified as a form of var. assamica. The leaves and young leaf buds of the native wild tea can also be used in the production of tea (Ho and Wang, 1984). Although they are not commonly exploited, the native wild teas provide an important reservoir of genes which can be introgressed into the current generation of clones. Their genetic relationship with cul
*Corresponding author. Tel: 886-4-22840417 ext. 315; Fax: 886-4-22874740; E-mail: jyhsiao@dragon.nchu.edu.tw