Bot. Bull. Acad. Sin. (2001) 42: 101-107
Hwu et al. Ribotoxin gene of Penicillium
Nucleotide sequence and the action of ribotoxin gene (sar gene) of Penicillium isolates from Taiwan
Luen Hwu1, Chiou-Jau Cho2, S. S. Tzeanz3, and Alan Lin2,*
1Institute of Microbiology and Immunology, National Yang-Ming University, Taipei 112, Taiwan, ROC
2Institute of Genetics, National Yang-Ming University, Shih-Pai, Taipei 112, Taiwan, ROC
3Department of Plant Pathology and Entomology, National Taiwan University, Taipei, Taiwan, ROC
(Received February 16, 2000; Accepted September 25, 2000)
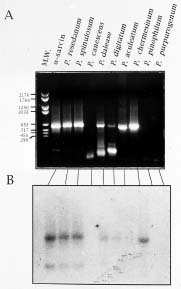
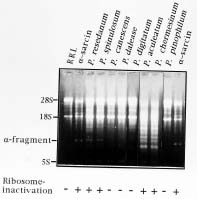
Abstract. Production of ribotoxin from species of Penicillium was reported. Among a total of fifty-six strains collected from Taiwan, six strains (P. resedanum, P. spinulosum, P. daleae, P. digitatum, P. aculeatum, and P. chermesinum) were found to carry ribotoxin gene (sar gene), but only four strains (P. resedanum, P. spinulosum, P. aculeaatum, and P. chermesinum) secreted the ribotoxic protein that cleaves ribosomes. The sar gene from the six strains was individually cloned and sequenced. These genes exhibit a typical genomic organization of the fungal-originated ribotoxin gene. The nucleotide and amino acid sequences of these sar genes from Penicillium are highly conserved and nearly identical. This is the first report that describes the production of ribotoxin by fungal species other than Aspergillus spp.
Keywords: Penicillium; Ribosome-inactivation; Ribotoxin; rRNA; sar gene.
Introduction
Filamentous fungi of Aspergillus and Penicillium are important in modern medicine and molecular biology, because they produce a wide range of secondary metabolites. Some of the metabolites have antibiotic or toxic properties in plants or animals. Aspergillus giganteus carries the sar gene that encodes a ribotoxic a-sarcin protein which inhibits protein synthesis of cells. The inhibition results from endonucleolytic cleavage of an RNA domain in 23-28S rRNA (Endo et al., 1983; Wool, 1984). One molecule of a-sarcin could catalyze the cleavage of 5000 molecules of ribosomes. This action has made ribotoxin a potential protein drug in the fight against cancer (Wawrzynczak et al., 1991; Brinkmann and Pastan, 1994; Lin et al., 1994).
Apparently, Aspergillus spp. is the only fungal species definitely known to carry the sar gene (Lin et al., 1995). Expression of the sar gene by several species of Aspergillus has been observed (Fernandez-Luna et al., 1985; Lamy and Davies, 1991; Lamy et al., 1991; Huang et al., 1997). The major human pathogenic species of Aspergillus is A. fumigatus; responsible for a variety of allergenic (Arruda et al., 1990) and invasive diseases (Vanden Bossche et al., 1988). The major cause of aspergillosis is the secretion of a ribotoxic protein by A. fumigatus (Lamy et al., 1991). The sar gene displays a unique genomic organization, in which an intron is inserted between exons of the signal peptide and the mature ribotoxin gene (Lamy and Davies, 1991;
Lamy et al., 1991; Huang et al., 1997). The origin and the need of the sar gene in Aspergillus have been questioned (Wool, 1984) since the gene is neither an essentially functional nor a house-keeping gene. Furthermore, expression of sar gene in cells can stop their growth or cause their death (Miller and Bodley, 1988; Lin et al., 1991; Hwu et al., 2000). This report describes findings of an investigation of possible production of ribotoxin in fungi other than Aspergillus spp. Species of Penicillium from collections in Taiwan that carried sar gene and secreted ribotoxic proteins were identified, and the DNA sequences of the genes were determined and compared.
Materials and Methods
Fungal Species and Culture Conditions
Species of Penicillium, including anamorphs and teleomorphs (Table 1), were obtained from the culture collections of Taiwan (Culture Collection and Research Center, Hsinchiu, Taiwan). All fungi were grown in 500 ml of potato dextrose broth (Difco Lab, Detroit, USA) in one-liter flasks. They were maintained at 30°C, with continuous agitation for 5 days. The mycelium were separated from the culture medium by filtering through Whatman No. 3 filter paper. Total DNA was extracted from the mycelium as follows: mycelium were frozen with liquid nitrogen and ground to a fine powder in a pre-chilled metal Waring blender cup. The powder was first suspended in a high salt containing buffer (1.5 M NaCl, 1.5 M EDTA, 100 mM Tris-HCl, pH 8.1), to which was added a 2% (the final concentration) cetyltrimethylammonium bromide (CTAB), reducing the salt concentration. The reduction
*Corresponding author. Tel: (02) 2822-5485; Fax: (02) 2826-4930; E-mail: alin@ym.edu.tw