Bot. Bull. Acad. Sin. (2001) 42: 115-121
Wu et al. Identification of Chlorella spp. isolates
Identification of Chlorella spp. isolates using ribosomal DNA sequences
Hsiuan-Lin Wu, Ruey-Shyang Hseu and Liang-Ping Lin*
Graduate Institute of Agricultural Chemistry, National Taiwan University, Taipei, Taiwan 106, Republic of China
(Received May 24, 2000; Accepted October 31, 2000)
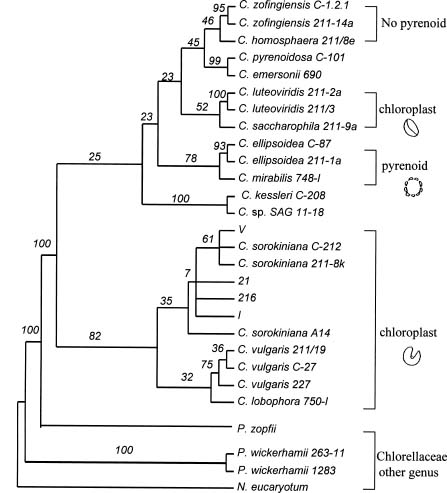
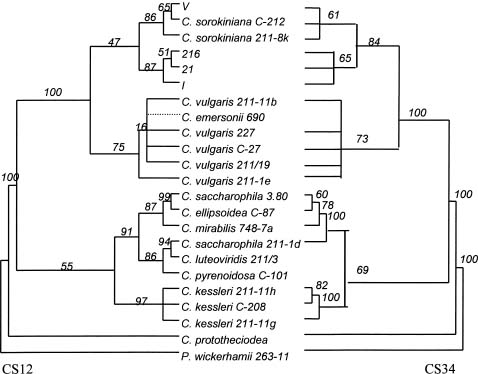
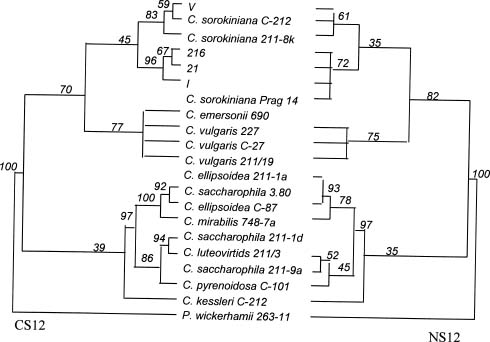
Abstract. Members of the Chlorella species are very simple unicellular algae, easy to cultivate and widely used in various physiological studies. Their morphological and physiological characteristics, however, normally change with the environment, making species identification difficult. To elucidate the relationship between various strains of Chlorella, this investigation analyzed the nuclear-encoded and chloroplast-encoded small-subunit rDNA sequences of four strains of Chlorella using PCR techniques. These strains were isolated from different rivers and ponds in Taiwan and Indonesia, and then compared and identified using stock strains of Chlorella spp. from the culture collection centers, and published DNA sequence data from Genbank. Experimental results attributed the isolated strains mainly to C. sorokiniana, a common species of green algae which grows in freshwater ecosystems at around 36°C. In addition, phylogenetic analysis of nuclear-encoded and chloroplast-encoded small-subunit rDNA sequences from spherical green algae of the genera Chlorella revealed the sequences to closely resemble each other. Further analyses indicated that Chlorella spp. 216 was close to Chlorella spp. 21 and I. Generally, the chloroplast data sets supported the lineages more than the nuclear data sets did. Strains 21 and 216 were closer to I. Comparisons with some of the morphological and biochemical data indicated that the phylogenetic analysis of rDNA sequences was in line with results obtained by conventional methods.
Keywords: Chlorella; Chloroplast; Nuclear; PCR; Phylogeny; Small subunit.
Introduction
The simple and common green algae of the genus Chlorella (Beijerinck, 1890) are placed below the order Chlorococcales and family Chlorellaceae (Hoek et al., 1995). Reproduction is asexual and achieved by producing non-motile autospores. Species of this genus are widespread in fresh water and in the sea, air, and soil. Warburg (1919) discovered that pure cultures of these fast-growing micro-organisms can be used as the ideal experimental materials for research on photosynthesis, nitrate reduction, physiology and biochemistry. Recently, Chlorella have been extensively studied and employed in various practical applications in agriculture and biotechnology. Chlorella are also used as protein-rich foods for sewage oxidation (Kessler, 1982). However, their cells do not exhibit characteristics that differentiate them from the morphological properties which are typically the basis of a classical taxonomic treatment of other algae (Shihira and Krauss, 1965).
Although the traditional taxonomic characteristics of Chlorella spp. indicate that morphological, biochemical and physiological properties are used in its identification, the cell size and shape are variable and largely depend on varying age, nutrition, and environmental factors (Fott and Novakova, 1969; Komárek and Fott, 1983). Additionally, certain biochemical and physiological characteristics are
not species specific (Kessler, 1982; Kessler, 1984; Kalina and Puncocharova, 1987). Therefore, classifying unknown isolated samples can be difficult.
The high number of copies and the inclusion of conserved and variable regions by evolution of ribosomal RNA genes (rDNA) have made possible a new method of species identification, classification and phylogenetic relationship determination (Mullis et al., 1986; Mullis and Faloona, 1987; Saiki et al., 1988). Algae cells have chloroplast genes just like higher plants, meaning cells have two varieties of SSU rDNA, nuclear-encoded and chloroplast-encoded. These characteristics have been considered for algae taxonomy and phylogenetic relationships (Huss and Sogin, 1990; Wilcox et al., 1992; Steinkötter et al., 1994; Schreiner, 1995). Meanwhile, Krienitz et al. (1996) compared the morphology and nuclear encoded SSU rDNA of green algae and found them closely related.
Recently, biochemical, physiological, and ultrastructural characters, together with molecular phylogeny based on the complete 18sRNA sequence, have led to the proposal that only four species should be kept in the genus Chlorella: C. vulgaris, C. lobophora, C. sorokiniana, C. kessleri (Huss et al., 1999).
Phylogenetic analysis of the order Chlamydomonadales revealed that this set of chloroplast data exhibited stronger support for comparable lineages than the set of nuclear data (Buchheim et al., 1996). Therefore, in this study field isolates from rivers in Indonesia and Taiwan are investigated, and the main observed cells were spherical,
*Corresponding author. Fax: 886-2-23626455; E-mail: m046@ccms.ntu.edu.tw