Bot. Bull. Acad. Sin. (2001) 42: 123-129
Chen et al. Sucrose phosphate synthase purification
Purification and characterization of sucrose phosphate synthase from sweet potato tuberous roots
Wei-Liang Chen1,3, Dong-Jiann Huang1,3, Pang-His Liu1, Heng-Long Wang2, Jong-Ching Su1,2, and Ping-Du Lee1,*
1Biochemistry Laboratory, Department of Agricultural Chemistry, National Taiwan University, 1 Roosevelt Road Section 4, Taipei, Taiwan 107, Republic of China
2Institute of Biological Chemistry, Academia Sinica, P.O. Box 23-106, Taiwan 107, Republic of China
(Received March 24, 2000; Accepted August 30, 2000)
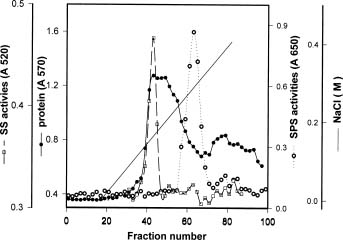
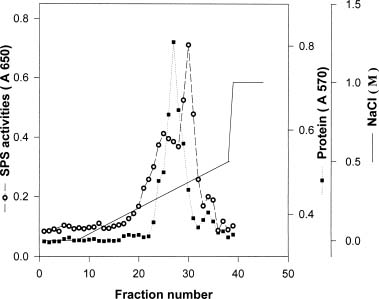
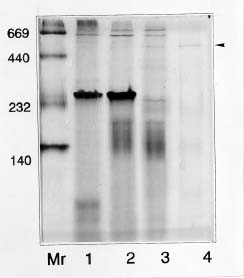
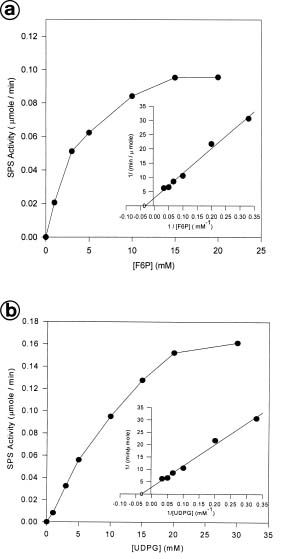
Abstract. Sucrose phosphate synthase (SPS) is one of the key enzymes in the sucrose biosynthesis pathway. SPS was purified 40 fold from crude extract of sweet potato tuberous roots by the methods of batch elution from DEAE-Sephacel, PEG precipitation, w-aminohexyl Sepharose 4B affinity and Mono Q anion exchange chomatographies. The native- and SDS-PAGE analyses revealed SPS to have a native molecular mass of about 540 kDa, and it may therefore be homotetramer composed of subunit with a mass of 130-140 kDa. The isoelectric point of the purified enzyme as determined by IEF was 5.29. SPS from the sweet potato tuberous root, which differs from the SPS of photosynthetic tissues, was not allosterically regulated by G6P and Pi. The Km for F6P and UDPG was 5.3 and 31.3 mM, respectively. The enzyme was activated by Mn2+, Mg2+, and Ca2+, while being inhibited by Hg2+. The nucleotides AMP, ADP, ATP, UMP, UDP, UTP, and TDP inhibited the enzyme about 30~50%. The enzyme was sensitive to sulfydryl reagents, but activity could be restored with DTT or b-ME. The enzyme was activated by glucose, glucosamine, maltose, and lactose, but was inhibited by d-gluconolactone. SPS could also be inhibited by PCMBS and Cibacron blue F3G-A.
Keywords: Enzyme purification; Ipomoea batatas; Sucrose metabolism; Sucrose phosphate synthase; Sweet potato.
Abbreviations: DTT, 1,4-dithiothreitol; b-ME, b-mercaptoethanol; PCMBS, p-chloromercuribenzenesulphonic acid; G6P, glucose 6-phosphate; F6P, fructose 6-phosphate; Pi, inorganic phosphate.
Introduction
Sucrose phosphate synthase (UDPglucose: D-fructose 6-phosphate 2-glucosyl-transferase; EC 2.4.1.14) catalyzes the following reaction:
UDPglucose + Fructose 6-phosphate « UDP + Sucrose 6'-phosphate
Sucrose phosphate synthase (SPS) catalyzes an essential and regulated reaction in the pathway of sucrose synthesis. In leaves, SPS is regulated by at least two mechanisms: one is an allosteric mechanism in which the enzyme is activated by binding metabolites like G6P and is inhibited by Pi (Doehlert and Huber, 1983), and the other one achieves regulation by covalent protein phosphorylation (Still et al., 1988).
In non-photosynthetic tissues such as tubers and fruits, SPS may play several important physiological roles, such as the biosynthesis and export of sucrose in germinating seeds and sprouting tubers, the accumulation of sucrose
under cold conditions, and the modulation of sucrose import and degradation via futile cycle in sink tissues. In non-photosynthetic tissues, the role of SPS is less well understood, and no obvious regulatory features have been found. To investigate the enzymatic function of SPS in sucrose biosynthesis, SPS has been purified to near homogeneity from spinach leaf (Salvucci et al., 1990), wheat germ (Salerno et al., 1991), maize leaf (Bruneau et al., 1991), Prosopis juliflora leaf (Sinha et al., 1997) and rice leaf (Salerno et al., 1998).
The objective of our work was to study the properties of the enzyme in detail and compare it with that of annual species. In this communication we report the purification and characterization of SPS from sweet potato tuberous roots.
Materials and Methods
Plant Material
Young, growing tubers of sweet potato, Ipomoea batatas (L.) Lam. cv. Tainoug 57, were harvested immediately before use.
3Both authors contributed equally to this work.
*Corresponding author. E-mail: pingdu@ccms.ntu.edu.tw