Bot. Bull. Acad. Sin. (2001) 42: 141-147
Ge and Chang Existence Analysis of Mosla hangchowensis
Existence analysis of populations of Mosla hangchowensis, an endangered plant
Ying Ge* and Jie Chang
College of Life Science, Xixi Campus, Zhejiang University, 232 Wensan Road, Hangzhou 310012, China
(Received December 13, 1999; Accepted September 4, 2000)
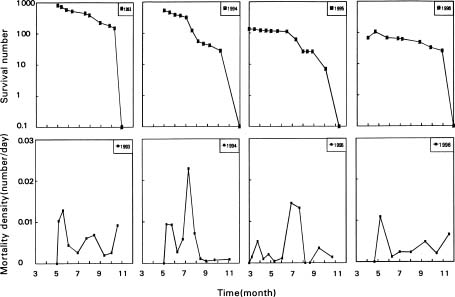
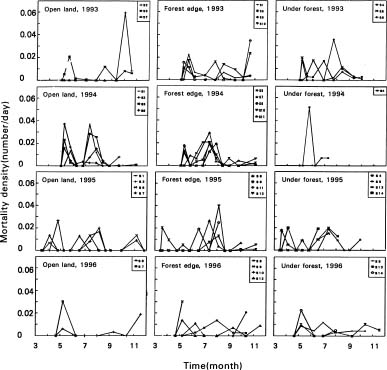
Abstract. Mosla hangchowensis Matsuch is an annual herb, which is found only on the seacoast of Zhejiang province in southeast China. The species has declined in numbers and is now endangered. The survival ratio, seminal power, reproductive success ratio, and density as well as other factors were measured from 1993 to 1996, and an existence analysis was performed. There are two environmental sieves during the life history of this species, and the survival curve is between the Deevey-I and Deevey-II. A constant proportion of individuals died throughout the life cycle. The optimum niche of M. hangchowensis is a habitat with soil depth 20-30 cm, with moderate to slightly wet conditions, with neither too intense nor too shaded light. The realized niche now is on the top of rocky hills or along gravel roadsides. Sites are few and dispersed. Loss of the optimum niche and a decrease of realized habitats as a result of human activities, the disparity between the realized niche and the optimum, and a low dispersal ability of populations are currently contributing to the endangerment of M. hangchowensis.
Keywords: Endangered; Existence analysis; Mosla hangchowensis; Population.
Introduction
Because of the rapid expansion of human activities, many species have fewer and fewer suitable habitats for their existence, and more and more species are becoming endangered or extinct. In recent years, many studies on conservation biology have been conducted all over the world in order to prevent biodiversity from disappearing. Not only have large-scale perennial species become endangered, many annual herbs have too (Hibig, 1982).
According to the basic principle of conservation biology, the survival ratio, seminal power, reproductive success ratio, population structure, species diversity, and density are the basic parameters that determine a species' degree of endangerment (Primack, 1993). If a low survival ratio was the primary cause of a species being endangered, then population trends and population survival analysis should be conducted (Feng, 1982) in order to find the time period in which high mortality occurred from the death density function curve. The environmental factors that caused death can be identified through a comparative analysis and used to define the environmental sieve that limits the growth of the population (Jiang, 1992).
Mosla hangchowensis Matsuch, an annual herb (0.2- 1.2 m in height), belonging to the mint family (Labiatae), has a small distribution area. There are only 5 small local populations extant along the coast in the eastern part of subtropical China, where it is endangered. The species is predominantly outcrossing and had a smaller seed-set
(77.5%) than did the congener, M. chinensis (95%), which is predominantly self-fertile and cosmopolitan (Zhou et al., 1996). Mosla hangchowensis possesses higher genetic diversity (Zhou et al., 1996) and lower seed germination (Ge et al., 1998a) than M. chinensis. In the study presented here, the survival of M. hangchowensis from young seedling to adult was analyzed, and the factors that caused the plant death were determined by a study of population ecology. Based on the trends in the growth and decline, we present the mechanism, leading factors, and key periods of population decline. Moreover, possibilities for remediation are discussed.
Study Sites
Mosla hangchowensis is distributed in island-shaped soil on big, bare rocks or along the sunny side of hill peaks. One local population can have three kinds of habitats:
(1) Open land habitat: There exist big bare rocks (from several to some ten square meter) with island-shaped soil covering them, the soil islands varying from ten square centimeters to several square meters, and the soil depth from 1-25 cm, 5-8 cm on average. There are two types of community: a. Mosla hangchowensis is the single dominant species in the community accompanied by only a few herbaceous and woody climber species, such as Commelina communis and Trachelospermum jasminoides living at the lower strata, the total biomass of which was only 3-5% of the total community. The population density of young plants in this habitat was high, varying from 100 to 2,400 individuals/m2. b. Mosla hangchowensis is not the dominant species; it intermixed with C. communis and T. jasminoides and some other species. In open land
*Corresponding author. Tel & Fax: +86-571-7972193; E-mail: jchang@mail.hz.zj.cn.