Bot. Bull. Acad. Sin. (2001) 42: 181-186
Turner et al. Molecular phylogeny of cyanobacteria
Molecular phylogeny of nitrogen-fixing unicellular cyanobacteria
Seán Turner1,3, Tan-Chi Huang2, and Shu-Miaw Chaw2,*
1Department of Biology, Indiana University, Bloomington, Indiana 47405-3700, USA
2Institute of Botany, Academia Sinica, Taipei 115, Taiwan
(Received August 25, 2000; Accepted December 11, 2000)
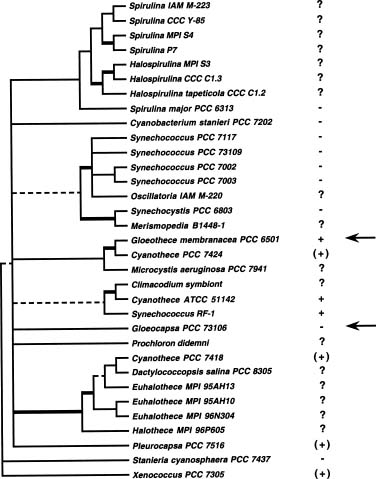
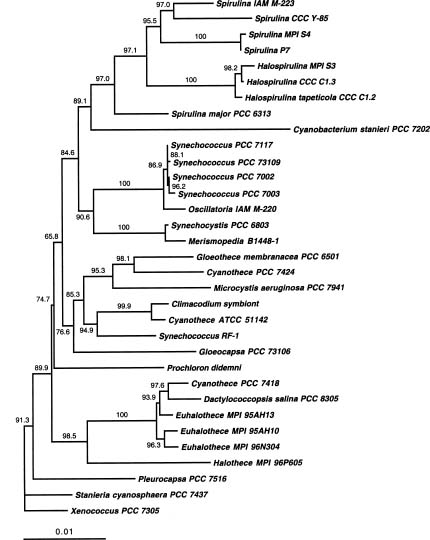
Abstract. Molecular phylogenetic study was conducted using maximum likelihood tree inference methods with small subunit ribosomal RNA sequence data to ascertain the evolutionary relationships among sheathless, single-cell cyanobacteria capable of nitrogen fixation. Cyanobacterial strains of the genus Cyanothece (circumscribed by Waterbury and Rippka, 1989) fall into at least three independent lines of descent within a larger assemblage previously designated the SPM sequence group. No strong correlation between aerobic versus anaerobic nitrogen-fixing activity and phylogenetic relationships was observed. The results support a hypothesis of multiple gains and/or losses of nitrogen-fixation abilities among the sheathless, unicellular cyanobacteria.
Keywords: Aerobic; Anaerobic; Cyanobacteria; Nitrogen fixation; Phylogeny; Ribosomal RNA; Sheathless; Small subunit.
Introduction
The unicellular cyanobacteria exhibit a great diversity of physiological properties. Few of them are able to fix nitrogen, either aerobically or anaerobically. Those capable of fixing nitrogen can be grouped into two types: sheathed and sheathless, the former being enclosed in a glycoprotein sheath (glycocalyx, capsule) external to the cell wall (Castenholz and Waterbury, 1989). The first sheathed species was classified in the genus Gloeocapsa (Wyatt and Silvey, 1969); however, it was later assigned to the genus Gloeothece, based on patterns of cell division (Rippka et al., 1979). On the other hand, the sheathless isolates were primarily attributed to the genus Synechococcus (Huang and Chow, 1986; León et al., 1986) or Aphanothece (Singh, 1973; Ni et al., 1988). More recently, Reddy et al. (1993) treated their sheathless isolates as members of Cyanothece, in agreement with the newer taxonomic criteria proposed by Waterbury and Rippka (1989).
Due to the fact that both botanical and bacteriological criteria have been used by various authorities to classify cyanobacteria, the systematics of these organisms has undergone numerous changes. (For a review of the problematic nature of cyanobacterial systematics, see Turner,
1997). The genus Cyanothece was first proposed by Komárek (1976) to accommodate some species previously placed in Synechococcus, the major feature distinguishing the former from the latter being that Cyanothece are present as single cells or in pairs, but never grouped into chains as are some Synechococcus. Subsequently, Waterbury and Rippka (1989) reserved the genus Cyanothece for unicellular cyanobacteria that lacked sheaths, divided in a single plane, and were larger than 3 µm in diameter. They also noted that, with one possible exception, the seven strains they originally placed in this genus were able to express nitrogenase activity either aerobically or anaerobically. The physiologies of these two kinds of nitrogen-fixing cyanobacteria are quite distinct. The aerobic type has developed a strategy to protect nitrogenase from oxygen generated by photosynthesis, but the anaerobic type either has not or has subsequently lost the ability to do so.
Because of the unique capability of fixing nitrogen and carbon dioxide within the same cell under aerobic conditions, the sheathless aerobic type has attracted more attention and has been better characterized than the anaerobic type (Mitsui et al., 1986; Huang et al., 1990; Colón-López et al., 1997). Therefore, our knowledge of the anaerobic type is very limited, and more studies are needed to justify whether the accommodation of the above two types of unicellular, nitrogen-fixing cyanobacteria in the same genus are reasonable. In this report, we present a phylogenetic analysis of small ribosomal subunit RNA (SSU rRNA) sequences to evaluate the phylogenetic relationships among sheathless, unicellular cyanobacteria with nitrogen-fixation abilities.
3 Present address: NCBI (GenBank), National Library of Medicine, NIH, Building 38A, 8600 Rockville Pike, Bethesda, Maryland 20894, USA.
*Corresponding author. Fax: 886-2-27827954; E-mail: bochaw@sinica.edu.tw