Bot. Bull. Acad. Sin. (2001) 42: 187-191
Sung and Chang Dormancy of azalea in Taiwan
Rhododendron mucronatum G. Don grown in subtropical Taiwan does not manifest endodormancy
F.H. Sung and Y.S. Chang*
Department of Horticulture, National Taiwan University, No. 1, Section 4, Roosevelt Road, Taipei 10617, Taiwan, Republic of China
(Received June 20, 2000; Accepted November 15, 2000)
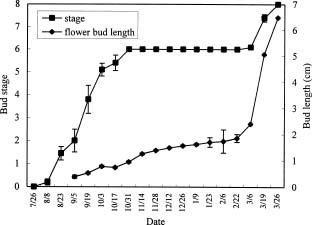
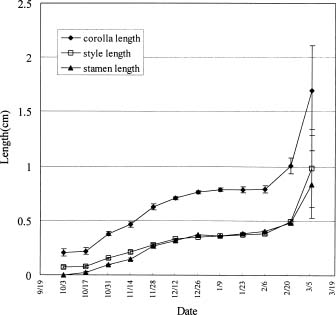
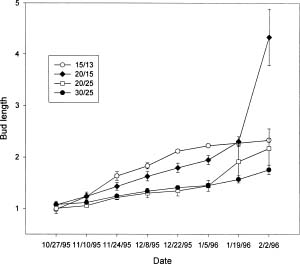
Abstract. Microscopic observation of bi-weekly sampled terminal buds revealed that most flower buds of R. mucronatum were formed between late-July and mid-August and differentiation of specific floral organs was completed before mid-October. Flower buds continued to enlarge through the winter. When flower buds on two-year-old plants from rooted cuttings were allowed to grow at day / night temperature of 15/13°C, 20/15°C, 25/20°C or 30/25°C, the 30/25°C treatment was observed to inhibit their growth; those subjected to the 15/13°C treatment grew the fastest during the early stages. Neither treatment, however, enabled attainment of anthesis by the end of the experiment. The 20/15°C treatment hastened flower bud growth and reduced the number of days to flowering. Bud growth was slower and flowering percentages were lower at 25/20°C, than at 20/15°C. When plants were transferred to 30/25°C after 2, 4, or 6 weeks in the 15/13°C treatment, the growth rate of flower buds increased greatly. Also, when the 15/13°C treatment was extended, the growth rate of flower buds increased, while the number of days needed for flowering after transferring from the low temperature treatment was reduced. The 15/13°C treatment simulated the winter temperatures in Taiwan. Our finding disclosed that bud growth was fastest, rather than ceasing, under this temperature regimen. Although growth rate decreased near the end of the experiment, it increased immediately and bloomed quickly when plants were transferred to higher temperatures. Thus, our observation indicated that R. mucronatum does not enter endodormancy during winter in subtropical Taiwan lowlands.
Keywords: Dormancy; Ecodormancy; Endodormancy; Flower bud development; Flowering; Temperature.
Introduction
Traditionally, it is believed that azalea flower buds enter dormancy before flowering and that the dormancy is overcome by low temperature. Hence, treatment of 4-6 weeks at 2-9°C is used to break flower bud dormancy for commercial production of potted azaleas (Larson, 1992; Seeley, 1981; Hamrick, 1991). When some azaleas, such as "Redwing" and "Reinhold Ambrosius," are grown at 20°C, they can attain anthesis without low temperature treatments; nevertheless, the uniformity of flowering is lower (Pettersen and Kristofferson, 1969). Brown and Box (1971) and Brown (1973) reported that some azalea cultivars, when grown at 18°C and under a 18-hour daylength, flowered about one-half to one month earlier than when given the traditional cold temperature treatment (5-week at 3°C); the uniformity of flowering was also very high. With temperate zone deciduous fruit species that are grown in tropical regions, Saure (1985) proposed that dormancy may be avoided because the warm temperature, together with the long daylength, is suitable for plant growth. This may be why some deciduous species can
bear fruits twice a year. Accordingly, the early flowering of some azalea cultivars as reported by Brown and Box (1971) and Brown (1973) may be the result of the warm temperature and long daylength having prevented dormancy from occurring in the flower buds. The relationship between temperature and azalea flower bud dormancy needs to be clarified more precisely.
Rododendron mucronatum is widely found in northern Taiwan lowlands. It was introduced in 1925, but there have been very few studies on its flowering habits and flower bud dormancy. Tang (1975) reported on its morphological changes, from vegetative growth to flower bud formation, but did not investigate flower bud dormancy. The purpose of our study has been to observe the flowering habit of R. mucronatum in Taiwan lowlands, and to evaluate the effect of temperature on flower bud development and dormancy. Our findings described here may help to control the flowering behavior of azalea in subtropical regions.
Materials and Methods
Determination of Flower Bud Developmental Stages
The flowering of ten azalea (R. mucronatum) plants, about 50-years-old and in good condition on the National Taiwan University campus, was observed from mid-July,
1Graduate student and associate professor of the Department of Horticulture, National Taiwan University.
*Corresponding author. Tel: 866-2-23630231-3340; Fax: 866-2-23635849; E-mail: yschang@ccms.ntu.edu.tw