Bot. Bull. Acad. Sin. (2001) 42: 207-214
Liu and Lin Ultrastructural study and lipid formation of Isochrysis sp.
Ultrastructural study and lipid formation of Isochrysis sp. CCMP1324
Ching-Piao Liu and Liang-Ping Lin*
Graduate Institute of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan, Republic of China
(Received May 22, 2000; Accepted December 12, 2000)
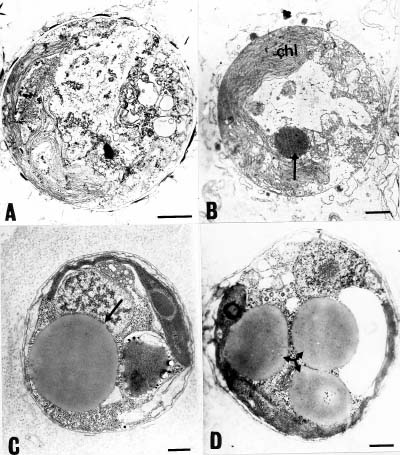
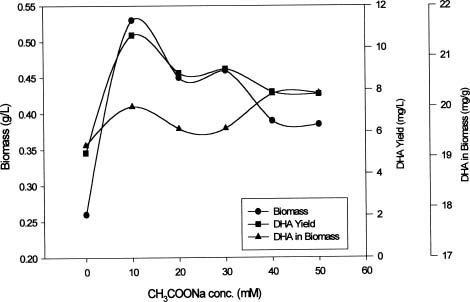
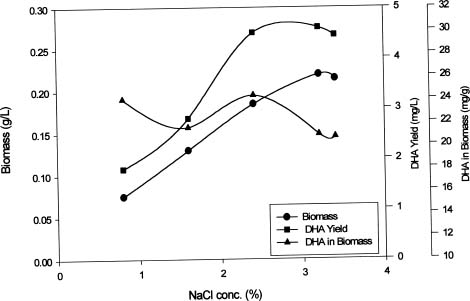
Abstract. This study investigates methods for extracting lipids from microalgae and analyzes the effects of culture media as well as culture conditions on PUFA yields and total fatty acid contents. Experimental results of an optimal culturing of Isochrysis spp. were based on a 3.2% salinity culture medium. These microalgae were cultured in a 1-2 L Roux's flat-flask and a 5 L jar fermentor. The optimum culture temperature and initial pH for DHA production were 25°C and 8.0, respectively. Pigments included chlorophylls a and c. The DHA yield increased with cultivation time until the eighth day. Optimum DHA amounts in the cells were reached under aeration with 10% CO2 and with continuous illumination of 10 klux. The biomass dry weight reached 4 g per liter of culture, and the DHA production reached 16 mg per liter of culture. Lipid bodies in Isochrysis spp. and related genera were observed during culture by light and transmission electron microscopy; 0.5~3.0 µm sized lipid bodies were confirmed by staining with Sudan Black B in cells from log stage to stationary stage cultures. These results demonstrated that DHA-containing lipid bodies in cells can be produced and accumulated in marine Isochrysis spp.
Keywords: Docosahexaenoic acid; Isochrysis sp.; Lipid formation; Polyunsaturated fatty acids (PUFA); Ultrastructure.
Introduction
Marine microalgae such as Isochrysis have received increasing interest because of their ability to produce the polyunsaturated fatty acid docosahexaenoic acid (DHA), one of the n-3 fatty acids believed to provide health benefits associated with the consumption of certain marine fish and their oils. DHA, a C22-polyunsaturated fatty acid, and its derivatives help prevent and treat pathologies such as coronary heart disease and atherosclerosis (Norday and Hansen, 1994), inflammatory problems, and some cancers, and are believed to play a role in infant nutrition (Conner and Neuringer, 1987). DHA accumulates in the membranes of nervous, visual, and reproductive tissues (Dratz and Deese, 1986). Polyunsaturated fatty acids are especially helpful in preventing heart and circulatory disease and facilitating brain development in infants (Yongmanitchai and Ward, 1991). Fish oils may not be an ideal source of n-3 PUFAs due to their scarcity and odor, as well as geographical and seasonal variations in quality (Varela et al., 1990).
Isochrysis has been widely used as a mariculture feed due to its high content of long chain polyunsaturated fatty acids (PUFAs) (Jeffrey et al., 1994). However, the lipid class and fatty acid compositions of microalgal cells at different growth phases can differ significantly (Emdadi and Berland, 1989), and can change with variations in culture conditions e.g. nutrient status, temperature, salinity, pH,
photoperiod, light intensity and light quality (reviewed by Yongmanitchai and Ward, 1989; Roessler, 1990). The cell structure of Isochrysis has attracted the attention of many investigators. Earlier studies (Green and Pienaar, 1977; Hori and Green, 1985; 1991) on Isochrysis galbana have mainly focused on its flagellar root system.
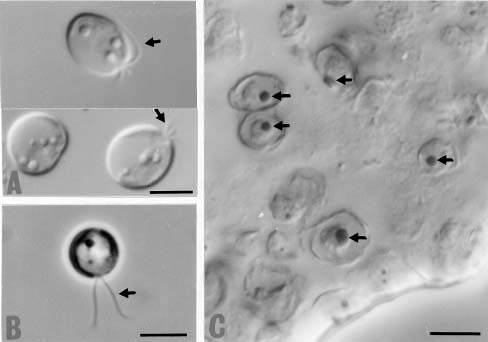
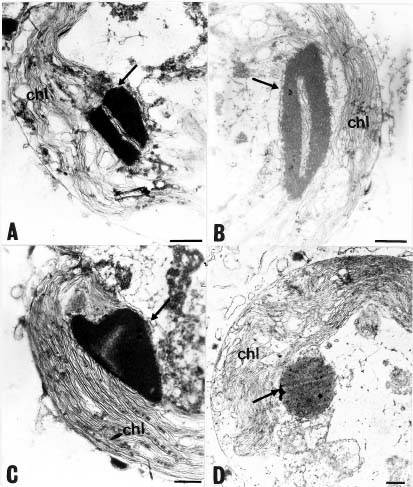
In this study, EM technologies were employed to survey this alga since previous papers have lacked detailed investigations of lipid formation in marine Isochrysis. This work also examines marine microalgae Isochrysis spp. as an alternative source of PUFAs and analyzes the culture medium and culture conditions that affect yields of PUFAs and their content in the total fatty acids. Lipid bodies in Isochrysis spp. and related genera are observed by light and transmission electron microscopy. Lipid granules are confirmed by staining with Sudan Black B in cells from the stationary cultures. The results suggest that DHA-containing lipid bodies in cells can be produced by marine Isochrysis. The possible commercial production of biomass and DHA-rich oil for use as food and feed ingredients is also predicted.
Materials and Methods
Cell Growth
Microalgal strains CCMP 463, 1324, 1325 and Pavlova salina were obtained from the Provasoli-Guillard Center for Culture of Marine Phytoplankton (West Boothbay Harbor, Maine USA). Isochrysis galbana TK1, TK2, were originally isolated by the Tungkang Marine Laboratory (Pingstung, Taiwan). Nannochloropsis oculata and Chlo
*Corresponding author. Fax: 886-2-23626455; E-mail: m046@ccms.ntu.edu.tw