Bot. Bull. Acad. Sin (2001) 42: 215-222
Chiang and Lin Diphyscium of Taiwan
Taxonomic revision and cladistic analysis of Diphyscium (Family Diphysciaceae) of Taiwan
Tzen-Yuh Chiangl,* and Shang-Hsiung Lin2
lDepartment of Biology, National Cheng-Kung University, Tainan 701, Taiwan
2Department of Biology, Tunghai University, Taichung 407, Taiwan
(Received April 2, 1999; Accepted September 7, 1999)
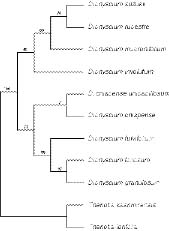
Abstract. Six species of Diphyscium, with three new additions to the moss flora of Taiwan, are revised taxonomically, including D. chiapense Norris ssp. unipapillosum (Deguchi) T. Y. Chiang and S. H. Lin, stat. nov. Diagnosable keys are provided. In order to test Norris' and Deguchi et al.'s classifications, cladistic analyses were carried out on eleven morphological traits scored from nine Asiatic species of Diphyscium. Two equally parsimonious trees rooted at the Theriotia species were recovered by PAUP. The reconstructed phylogeny, which suggested a close relationship between the taxa with single papillose lamina cells and the species with pluripapillose lamina cells, invalidated Norris' hypothesis. The parsimony tree agrees with Deguchi et al.'s classification, in which three groups were classified based on cell-wall papillosity. Based on the inferred phylogeny, the "orifice" stomata may have evolved more than once in Diphyscium, while a single "secondary loss" event occurred in the group of D. involutum, D. rupestre, D. mucronifolium and D. suzukii.
Keywords: Cladistics; Diphyscium; D. chiapense ssp. unipapillosum; Orifice stomata; Papillosity; Phylogeny; Taiwan; Taxonomy.
Introduction
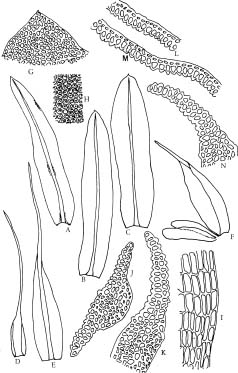
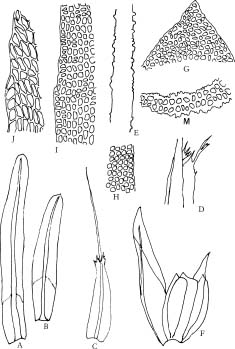
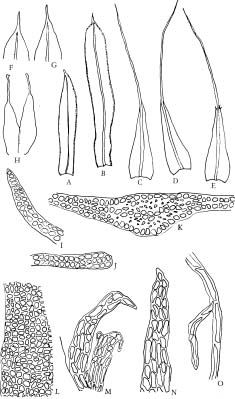
Diphyscium is a conspicuous moss characterized by its obliquely ovoid capsule with dorsiventral differentiation and a narrow mouth as well as strongly differential, bristled perichaetial leaves. Diphyscium and Theriotia constitute the Diphysciaceae. Diphyscium is distinguished from the latter by the well differentiated laminate portion of the leaves, waxy anticlinal walls in the dorsal side of the capsules, and the absence of chlorophyllosd cells in the leaves (cf. Deguchi, 1984a). Phylogenetically, Diphysciaceae are closely related to the Buxbaumiaceae, sharing common structure and ontogeny of the capsules (Crum and Anderson, 1981). The reduced gametophytes and elongated setae in Buxbaumiaceae, however, suggest an independent lineage from the Diphysciaceae. Despite the remarkable differences of the sporophytic structure, the gametophytes of Diphyscium in some cases [e.g., D. pocsii (Bizot) Zand.] resemble the Pottiaceae. Deguchi (1975, 1984b) and Norris (1981) utilized the "collared" axillary hairs to differentiate the Diphysciaceae from the Pottiaceae.
About twenty-three species have been recognized, with a distribution ranging from the tropics to the temperate (van der Wijk et al., 1967; Crum and Anderson, 1981;
Crosby et al., 1992; Crosby and Magill, 1994; Deguchi et al., 1997). Deguchi et al. (1997) distinguished three groups of Diphyscium based on the papillosity of their leaf-cell surface: 1) species with smooth leaf cells (e.g., D. involutum), 2) species with unipapillose-mamillose leaf cells (e.g., D. chiapense), and 3) species with pluripapillose-mamillose leaf cells (e.g., D. fulvifolium). In contrast, Norris (1981) recognized two groups, with D. foliosum and D. involutum being representative. Diphyscium chiapense was regarded as a member of the D. involutum group according to his classification. Although Deguchi et al. (1997) treated D. chiapense and its relatives as a new group, the phylogenetic relationships among the three groups were not further discussed and therefore have remained unknown.
Interestingly, although the plants have discernible morphological characteristics, little research has been carried out on the east Asiatic (i.e. As 2; van der Wijk et al., 1967) taxa except for the Japanese species (Iwatsuki, 1976; Deguchi, 1983, 1984b; Noguchi, 1987; Deguchi et al., 1997). For example, the floristic survey of the Chinese Diphyscium has been poorly documented, with only four species and one variety known (cf. Redfearn et al., 1996). The tiny plants, with pottiaceous appearance when the sporophytes are absent, may have caused it to be overlooked by collectors.
The first record of this genus in Taiwan is from 1935, with the collections of Diphyscium fulvifolium and D. formosicum (= D. involutum) (Horikawa, 1935a, 1935b).
*Corresponding author. Tel: +886-6-2757575 ext. 65525; Fax: +886-6-2742583; E-mail: tychiang@mail.ncku.edu.tw