Bot. Bull. Acad. Sin. (2001) 42: 231-241
Fu et al. Patatin promoter-like sequence in Forever Young genes
Potato and tomato Forever Young genes contain class-I patatin promoter-like sequences
Hongyong Fu1,*, Ji Du2,4, Junqi Song3, Jiming Jiang3, and William D. Park2,*
1Institute of Botany, Academia Sinica, 128, Sec 2, Academy Rd., Nankang, Taipei, Taiwan 11529, Republic of China
2Department of Biochemistry and Biophysics, Texas A & M University, College Station, Texas 77843, USA
3Department of Horticulture, University of Wisconsin, Madison, Wisconsin 53706, USA
(Received December 21, 2000; Accepted July 3, 2001)
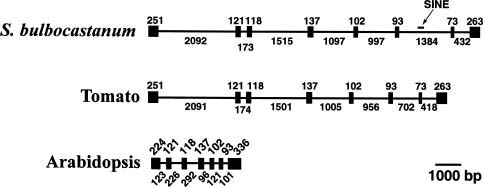
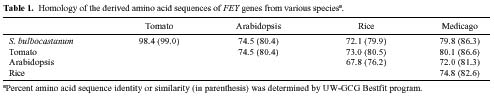
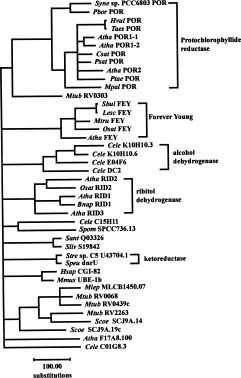
Abstract. Class-I patatin genes from potato, encoding the major storage protein in tubers, contain promoter repeat sequences, which are critical for developmental and metabolic regulation. Previously, RFLP mapping indicated the patatin genes of potato and tomato reside in orthologous loci near the end of the long arm of chromosome VIII. Interestingly, a DNA sequence homologous to the promoter of the potato class-I patatin gene was mapped to the middle of chromosome III in the tomato and potato genomes [Ganal et al., Mol. Gen. Genet. 225, 501-509 (1991)]. We isolated and characterized genomic clones containing the class-I Patatin Promoter-Like sequences (PPL) from tomato, S. bulbocastanum, and S. tuberosum. Sequence analyses reveal that these PPL sequences are highly conserved, as compared to the repeat sequence of the patatin promoter, particularly with respect to the critical regulatory cis-elements. These PPL sequences are located upstream of the translational initiation site of Forever Young genes, which encode proteins belonging to a large family of oxidoreductases with conserved structural motifs. The Forever Young genes from tomato and S. bulbocastanum both contain eight exons and seven introns. Six of the introns are uniquely large, in the range of 418~2,092 bp. Sequence conservation suggests that the class-I patatin repeat and the PPL from Forever Young genes may have a direct phylogenetic relationship and may regulate expression of their respective genes through common cis- and trans-factors.
Keywords: Forever Young; Oxidoreductase; Patatin; Potato; Promoter evolution; Tomato.
Introduction
Patatin is a family of glycoproteins that have lipid acyl hydrolase and transferase activities (Andrews et al., 1988) and account for up to 40% of the total soluble protein in potato tubers (Paiva et al., 1983). In potato, the patatins are encoded by a gene family with ~10-15 members per haploid genome. These genes can be divided in about equal numbers into two classes, class-I and class-II, based on the presence or absence of a 22-bp insertion in the 5´ UTR. Whereas these two classes of genes have conserved coding regions, the 5´ flanking sequences are highly diverged (Pikaard et al., 1987; Mignery et al., 1988). The class-I genes encode the majority of tuber patatins and have an essentially tuber-specific expression pattern
(Paiva et al., 1983). Besides being regulated developmentally, the class-I genes are also regulated by metabolic signal(s) derived from sucrose (Paiva et al., 1983; Rocha-Sosa et al., 1989; Wenzler et al., 1989; Jefferson et al., 1990). In contrast, patatins detected in roots and anther epidermal cells are primarily derived from class-II genes (Pikaard et al., 1987; Mignery et al., 1988). Unlike class-I genes, class-II genes are not sucrose-inducible (Köster-Töpfer et al., 1989; Liu et al., 1991).
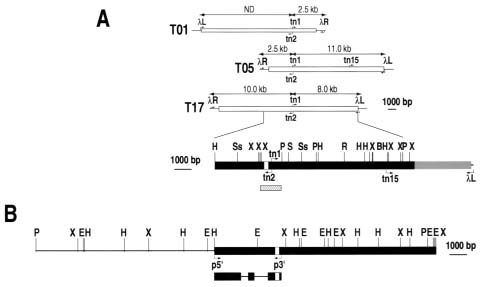
Cis- and trans-factors responsible for developmental and metabolic regulation of the class-I patatin genes from potato have been examined extensively. An imperfect repeat in the 5´ region of the class-I patatin promoter contains separable cis-acting elements that are critical for "tuber-specific" and sucrose-inducible regulation. The sequence can be divided into two sub-regions (box A and box B), which are positioned from -552 to -514 and -626 to -592, respectively, for the distal repeat (in referring to the transcription start site as +1; Mignery et al., 1988). Motifs within these repeat sequences were found to interact with nuclear factors (Jefferson et al., 1990; Liu et al., 1990; Grierson et al., 1994).
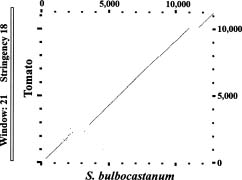
Genetic and physical mapping analyses indicated the patatin genes show syntenic chromosomal organization in potato and tomato genomes and were mapped as a single cluster of loci near the end of the long arm of chromo
4Current address: Emory University, Atlanta, GA 30322, USA.
5The amino acid and nucleotide sequence data reported for S. bulbocastanum FEY, S. tuberosum FEY, Tomato FEY, and Arabidopsis FEY3, will appear in the GenBank, EMBL and DDBJ databases under the accession numbers AF216836, AF250031, AF216835 and AF217275, respectively.
*Corresponding authors. H. Fu, Tel: 886-2-27823050 ext 422, Fax: 886-2-27827954, E-mail: hongyong@gate.sinica.edu.tw; W.D. Park, Tel: 979-845-8868, Fax: 979-862-4471, E-mail: wdpark@tamu.edu