Bot. Bull. Acad. Sin. (2001) 42: 273-279
Lin et al. Centromere position in maize
RFLP mapping of the centromere of chromosomes 1, 6 and 9 by B-A translocations in maize
Bor-Yaw Lin*, S.-J. Chang, and H.-M. Lin
Institute of Molecular Biology, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
(Received October 16, 2000; Accepted June 27, 2001)
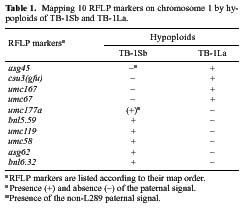
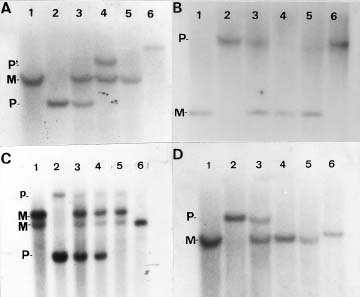
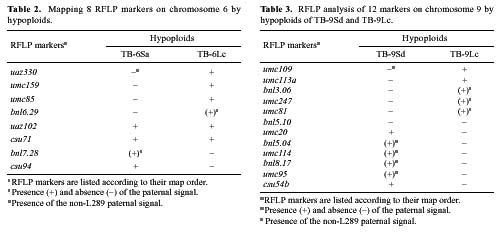
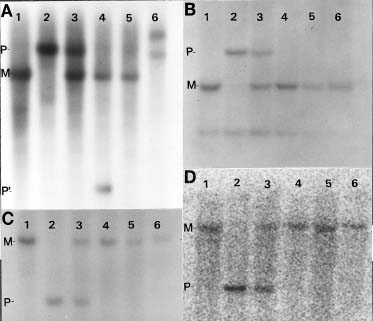
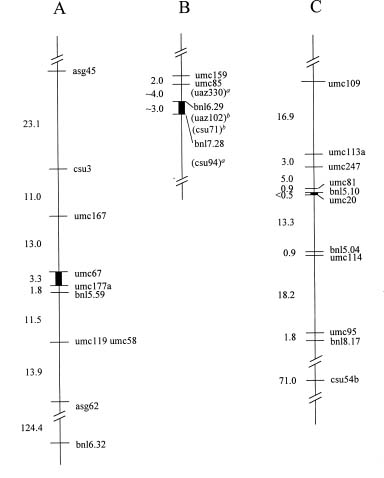
Abstract. The centromere of chromosomes 1, 6 and 9 are physically mapped by the hypoploids of the six most proximal B-A translocations. The hypoploids are deficient for a paternal chromosome arm and, as a result, lose the paternal signal of those RFLP markers located on the missing chromosome arm. Of those markers missing from the hypoploids, the two most proximal ones on each arm of a chromosome define the physical location of the centromere. Analysis of 10 RFLP markers on chromosome 1, 8 on chromosome 6 and 12 on chromosome 9, maps the first centromere to the umc67-umc177a region, the sixth centromere to the bnl6.29-bnl7.28 region, and the ninth centromere to the bnl5.10-umc20 region, an interval of about 3.3, 3 and 0.5 map units, respectively. Other interesting observations are that the A-B chromosome of five of the six B-A translocations is associated with anomalous signals not originated from the paternal parent, suggesting probable presence of chromosome rearrangement(s).
Keywords: B-A translocation; Centromere; Deletion mapping; Hypoploid; RFLP marker.
Introduction
The centromere is a distinct cytological regionthe primary constrictionon metaphase chromosomes. Its primary function is to align chromosomes at the metaphase plate and then to separate them at anaphase during cell division. It exerts these functions through the attachment of its kinetochore with microtubules, to promote chromosome movement by the kinetochore motor (Hyman and Mitchison, 1991). Despite its significant biological function, the centromeric location is rarely determined and has been mapped only in a few species due to lack of a proper phenotype for genetic analysis. This stumbling block was circumvented in yeast (Clarke, 1990) and recently in Arabidopsis (Copenhaver et al., 1999) using tetrad analysis to define the centromere position at the molecular level.
In plants, the centromere has been mapped with a telocentric or an isochromosome. The telocentric chromosome is derived from a normal homolog, usually a univalent, by a misdivision of the centromere; semicolon and one end of this telocentric chromosome is the centromere. An isochromosome is structurally similar to a telocentric. It is a metacentric chromosome, carrying two identical chromosome arms. Any genetic marker located at or near the terminal position on a telocentric will be either most proximal or distal to the centromere. After comparing with the genetic map of the normal homolog, the centromeric position can be determined to be located between two closely
linked genetic markers. Centromeres have been mapped this way in wheat (Sears, 1969), cotton (Endrizzi and Kohel, 1966), rice (Singh et al., 1996) and tomato (Frary et al., 1996). Telosomes associated with chromosomes 4 and 5 have been isolated in maize (Rhoades, 1933, 1936 and 1940; Muzumdar et al., 1997) and used to map the centromere of chromosome 4 (Schneerman et al., 1998).
In addition to telocentric chromosomes, a maize centromere has been well localized by hypoploids generated from B-10 translocations. Beckett (1973) induced a translocation between the B chromosome and the short arm of chromosome 10 (10S). He used the translocation to synthesize hypoploids carrying the terminal deficiency of 10S to map physically the 10th centromere to be proximal to y9 on 10S. Subsequently, Lin (1974) created 38 translocations between the B chromosome and the long arm of chromosome 10 (10L) and observed that the hypoploids derived from two most proximal translocations have a breakpoint proximal to du1 on 10L (Lin, 1974 and 1979). Thus, the 10th centromere is located in the du1-y9 region which covers two map units.
This paper utilizes a similar approach to map the centromere of chromosomes 1, 6, and 9 on RFLP map. The centromere was mapped to a region of 3.3, 3, and 0.5 map units, respectively.
Materials and Methods
Plant Material
L289 and the B-A translocation-carrying L289 stocks are the same as previously described (Lin and Chou, 1997). Three translocations (TB-1Sb, TB-1La and TB-6Sa), iso
*Corresponding author. Fax: (04) 2287-4879; E-mail: bylin@dragon.nchu.edu.tw