Bot. Bull. Acad. Sin. (2001) 42: 281-286
Yu et al. Effects of carbenicillin and cefotaxime on papaya regeneration
Effects of carbenicillin and cefotaxime on callus growth and somatic embryogenesis from adventitious roots of papaya
Tsong-Ann Yu1, Shyi-Dong Yeh2, and Jiu-Sherng Yang1,*
1Department of Botany and 2Department of Plant Pathology, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
(Received November 6, 2000; Accepted May 24, 2001)
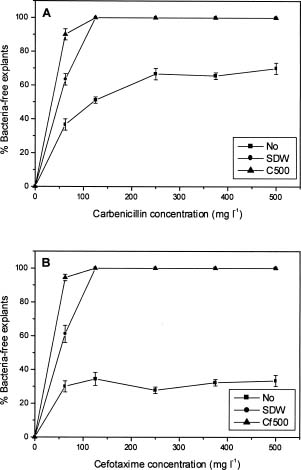
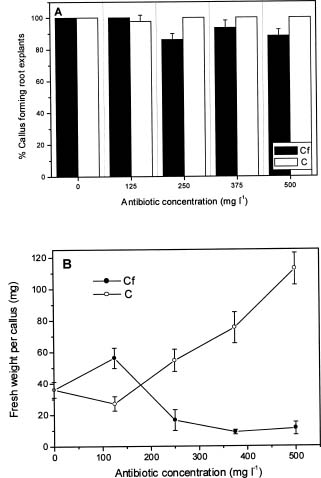
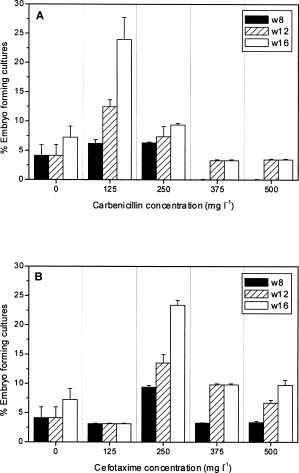
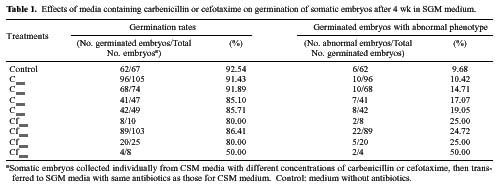
Abstract. Carbenicillin and cefotaxime, two antibiotics commonly used for excluding Agrobacterium tumefaciens during plant transformation, were tested for their bacteriostatic effects as well as for their effects on plant regeneration in adventitious root explants of papaya following co-culture with Agrobacterium. A washing step with sterilized distilled water two days after co-culture enhanced the bacteria-suppressing effects of antibiotics. Proliferation of Agrobacterium was completely suppressed in the medium containing 125 mgl-1 carbenicillin or cefotaxime. Callus fresh weight increase was apparently enhanced in the media with higher concentrations of carbenicillin (250-500 mgl-1), but was extremely inhibited in media with the same concentrations of cefotaxime. Higher percentages of somatic embryos were found in the medium with 125 mgl-1 carbenicillin or 250 mgl-1 cefotaxime; however larger numbers of somatic embryos from the individual callus were obtained in the medium with 125 mgl-1 carbenicillin than in the medium with 250 mgl-1 cefotaxime. Percentages of abnormal somatic embryos were lower in the medium with lower concentrations of carbenicillin (125-250 mgl-1). Favorable conditions for use of the two antibiotics for suppressing bacteria growth and enhancing regeneration of papaya plantlets from adventitious roots are discussed.
Keywords: Antibiotics; Bacteriostatic effect; Somatic embryogenesis.
Introduction
Considerable progress has been made in plant transformation during the last decade, using in many instances Agrobacterium mediation (Nauerby et al., 1997). For Agrobacterium-mediated DNA transfer, antibiotic use in plant tissue culture has become routine. Antibiotics are added to culture media to control Agrobacterium that may affect the plant regeneration process and to select transformants with an antibiotic-resistance that is cotransferred with the gene of interest (Shaw et al., 1983). Although many antibiotics have been described for effective control of Agrobacterium cells, carbenicillin and cefotaxime, both belonging to the b-lactam group, have minimal toxicity on most plant tissues (Mathias and Boyd, 1986) and thus have become most widely accepted in Agrobacterium-mediated transformation. However, both antibiotics have been known to have plant hormone-like effects on cultured plant tissues and could affect somatic embryogenesis in many plant species (Nauerby et al., 1997).
For Agrobacterium mediated gene transfer in papaya, carbenicillin (Fitch et al., 1990; 1993; Yang et al., 1996; Cheng et al., 1996) and cefotaxime (Fitch et al., 1993; Cabrera-Ponce et al., 1996) are often added to the medium during plant regeneration to control Agrobacterium growth, but information about their effects on develop
ment of callus and somatic embrys of papaya has not been considered.
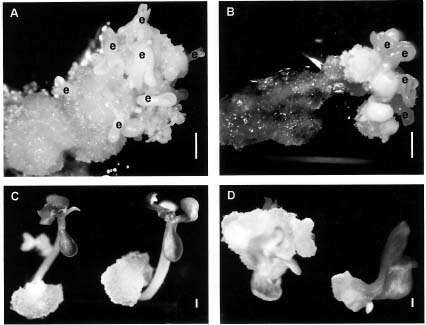
Immature embryos are commonly used for papaya transformation (Fitch et al., 1990; Cheng et al., 1996). However, their use is time consuming and hindered by a seasonal factor. Moreover, the desired sex and other cultivar traits of transgenic lines can only be determined several months after planting. Recently, we established an efficient system for inducing adventitious roots from in vitro shoots, derived from selected hermaphrodite plants with good horticultural qualities (Yu et al., 2000). The adventitious roots thus obtained can regenerate somatic embryos within 4 months. Regeneration from adventitious roots avoids the disadvantages of immature embryos. However the parameters for Agrobacterium-mediated transformation using adventitious roots have remained unestablished. In this investigation, root segments were co-cultured with Agrobacterium, then treated with carbenicillin and cefotaxime to investigate the antibiotic effects. Also, the effects of carbenicillin and cefotaxime on induction of callus, initiation of somatic embryos, and germination of somatic embryos during the plant regeneration process from papaya root segments were investigated.
Materials and Methods
Plant Materials and Culture Conditions
Source of adventitious roots: Individual shoots (>5 mm) with 2-3 leaves from multiple shoots of papaya (Carica papaya L. cv. Tainung No. 2) that arose in vitro
*Corresponding author. Tel: +886-4-2840417 ext. 404; Fax: +886-4-2874740; E-mail: jsyang@dragon.nchu.edu.tw