Bot. Bull. Acad. Sin. (2002) 43: 1-11
Ashikari and Matsuoka Application of rice genomics to plant biology and breeding
(Invited review paper)
Application of rice genomics to plant biology and breeding
Motoyuki Ashikari* and Makoto Matsuoka
Bioscience Center, Nagoya University, Chikusa, 464-8601, Japan
Keywords: Brassinosteroid; Dwarf; Genetics; Genomics; Gibberellin; Marker assisted selection; Molecular biology; Molecular breeding; Quantitative trait loci; Rice.
Abbreviations: RFLP, Restriction Fragment Length Polymorphism; GA, Gibberellin; BR, Brassinosteroid; MAS, Marker Assisted Selection; QTL, Quantitative Trait Loci; KD, Kinase Domain.
Contents
Rice Genomics 1
Application to Biology 3
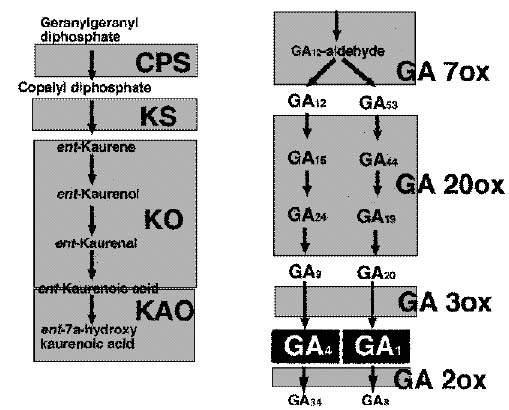
GA B0iosynthetic Pathway 3
GA Signal Transduction Pathway 4
d1 mutant 4
slender (slr) Mutant 4
Isolation of New Genes Associated with GA Signaling 5
gibberellin insensitive dwarf 1(gid1) Mutant 5
Application to Breeding 5
Molecular Breeding 6
Improvement of Plant Height by Controlling GA-Associated Genes 6
Improvement of Rice Plant Architecture by Expression of the Dominant Negative OsBRI1 7
Marker Assisted Breeding 8
QTL Approach 8
Future Outlook 9
Literature Cited 9
Rice Genomics
It has been estimated that 50% of the human population depends on rice (Oryza sativa L.) as its main source of nutrition (White, 1994). In particular, it is the most important crop for people living in the monsoonal areas of Asia, where it has a long history of cultivation and is deeply ingrained in the daily lives of Asian people.
For many years, rice has been the subject of numerous breeding studies aimed at developing higher yielding or better tasting cultivars. It has also become a useful plant for studying biology, as a model plant of monocotyledons (monocots). Rice has a small genome (430 Mb) relative to other Gramineae plants, and its genome size is about three times larger than that of Arabidopsis, a model plant of dicotyledons (dicots). Last decade, technological innovations in science enabled dramatic advancements in the
field of plant genomics (genome science). Many rice genome projects have been launched, and they are providing very useful information for plant biology and plant breeding.
Between 1966 and 1990 the population of densely populated and low-income countries grew by 1.8 fold while food production more than doubled. Such a rapid increase in the volume of world food production was mainly due to the development of high-yielding varieties of wheat and rice and other technological advancements (Khush, 1999). Now, however, the rate of world population growth has exceeded the rate of growth in food-grain production. It is predicted that the world population will exceed 8 billion people by 2025, with the greatest increase occurring in developing countries. Actually, many people living in Asia and Africa already suffer from malnutrition. To meet these global food demands, grain production will need to increase 50% by 2025 (Khush, 1999). Prompt measures and action should be taken to avoid a large famine and widespread food scarcity.
*Corresponding author. Tel: 81-85-789-5225; Fax: 81-52-789-5226; E-mail: ashi@nuagr1.agr.nagoya-u.ac.jp