Bot. Bull. Acad. Sin. (2002) 43: 37-56
Tsou and Mori Seed coat of Lecythidoideae
Seed coat anatomy and its relationship to seed dispersal in subfamily Lecythidoideae of the Lecythidaceae (The Brazil Nut Family)
Chih-Hua Tsou1 and Scott A. Mori2,*
1Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
2Nathaniel Lord Britton Curator of Botany, Institute of Systematic Botany, The New York Botanical Garden, Bronx, New York 10458-5126, USA
(Received April 19, 2001; Accepted August 31, 2001)
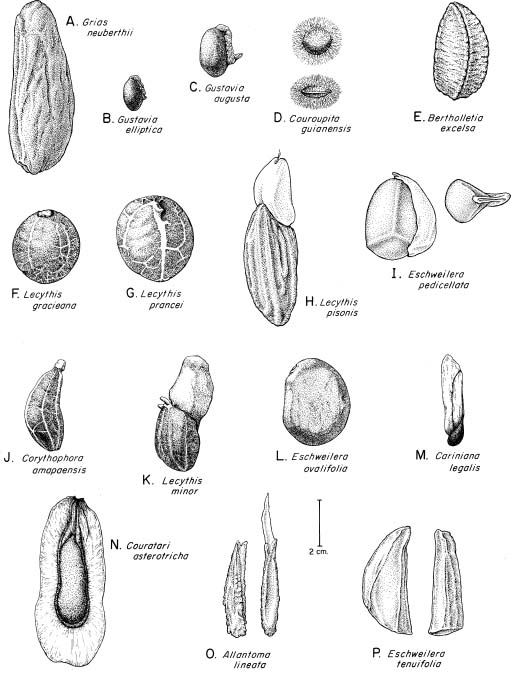
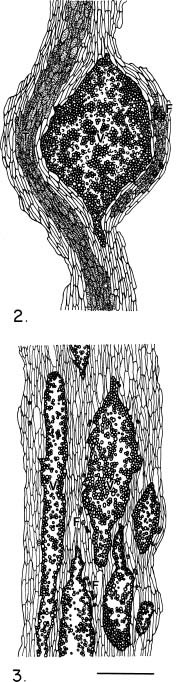
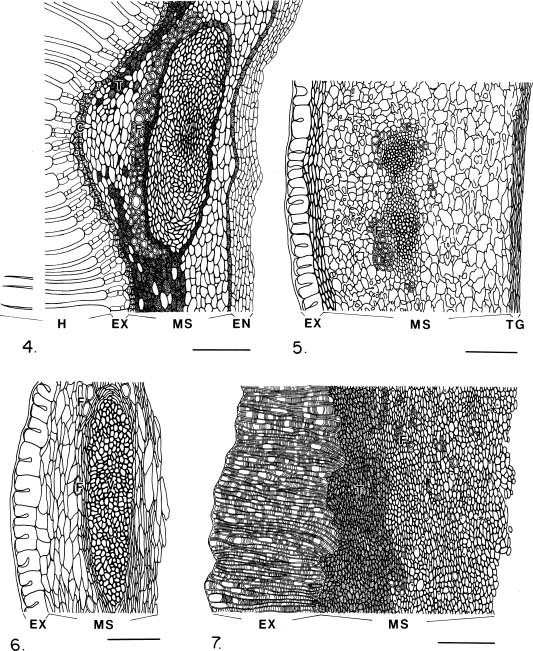
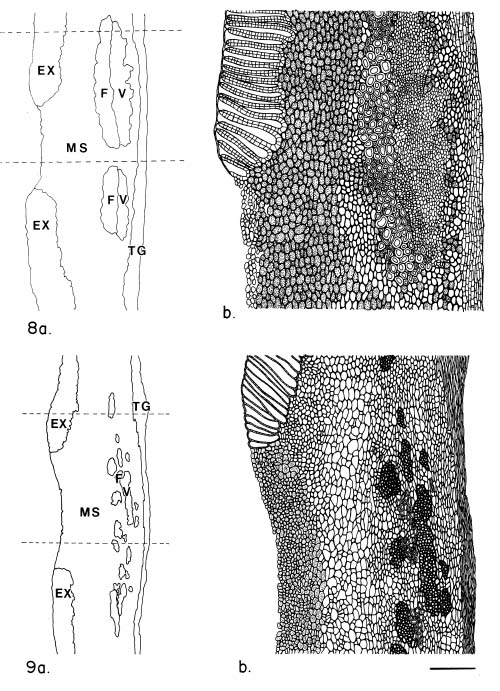
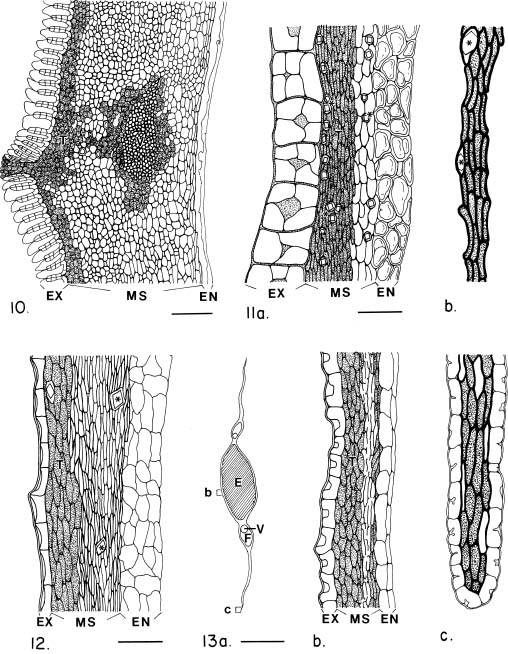
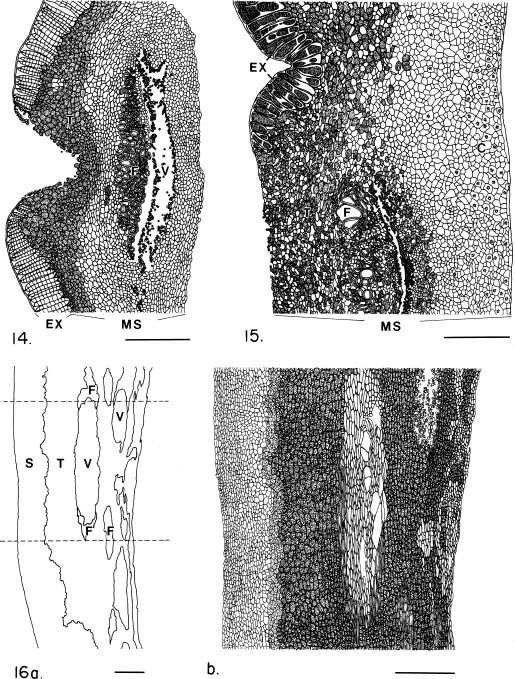
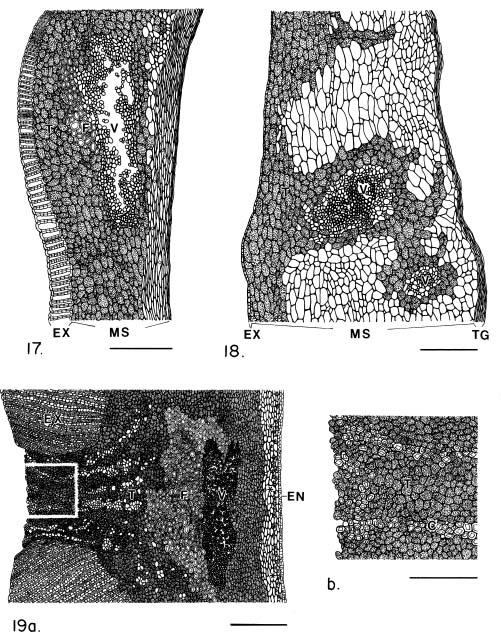
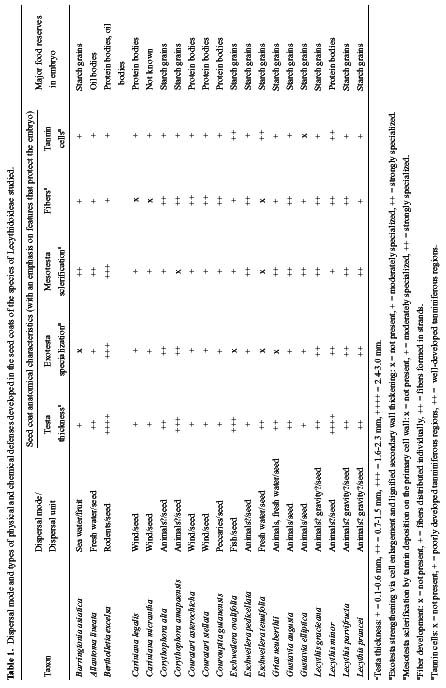
Abstract. The seed coat anatomy of representative species from all 10 Neotropical genera of Lecythidaceae subfamily Lecythidoideae and from the Paleotropical Barringtonia (Lecythidaceae subfamily Planchonioideae) was studied. The seed coat is mainly composed of the testa, which is developed through moderate or intensive multiplication of the outer integument of the ovule. The tegmen, derived from the inner integument of the ovule, is mostly crushed at seed maturity. Barringtonia and Grias, with fruits as diaspores, have an unspecialized exotesta and a poorly differentiated seed coat. In contrast, species of Lecythidoideae, with seeds as diaspores, possess well-differentiated seed coats with diversified protective mechanisms. Examples include: an expanded and lignified exotesta that serves as a water barrier and protects the embryo; an extensive area of tannin cells that provides a chemical defense against pathogens and predators; a thick and sclerotic mesotesta that protects the embryo; and large fibers surrounding and supporting the vascular bundles. In addition, other modifications of the seed coat are probably adaptations for seed dispersal by different dispersal agents. These include the exotestal hairs of Couroupita guianensis, which possibly protect the embryos as they pass through the digestive tracts of peccaries; an extension of the seed coat into a wing in species of Cariniana and Couratari , which facilitate wind dispersal; the papillate exostestal cells of Allantoma lineata, which probably promote dispersal by water; the well-developed arils of species of Corythophora and Lecythis, probably sought after by animals, which in turn disperse the seeds; and the sarcotesta of Eschweilera ovalifolia, consumed by fish, which in turn disperse the seeds. We conclude that the highly diversified anatomy and morphology of the seed coat of the Lecythidoideae has evolved from relatively homogeneous ovules in response to pathogens and predators as well as to abiotic and biotic dispersal agents.
Keywords: Brazil nut family; Lecythidaceae; Lecythidoideae; Planchonioideae; Seed coat anatomy; Seed dispersal.
Introduction
The Brazil nut family (Lecythidaceae), a pantropical family of small to very large trees, is represented by 11 genera and approximately 200 species in the New World tropics. All New World Lecythidaceae, except Asteranthos brasiliensis Desf. (Lecythidaceae subfamily Scytopetaloideae), belong to Lecythidaceae subfamily Lecythidoideae (Prance and Mori, 1979; Mori and Prance, 1990a; Morton et al., 1997, 1998). Barringtonia is an Old World member of Lecythidaceae subfamily Planchonioideae and is sister to the Lecythidoideae (Tsou, 1994).
New World Lecythidaceae are an important element of the flora at latitudes between 19° N and 25° S (Mori and Prance, 1990a). They rank as the first or second most important family of trees in terms of species and numbers of individuals in the vicinity of Belém, Brazil (Black et al., 1950;
Cain et al., 1956). They are the third most important tree family at La Fumée Mountain, French Guiana (Mori and Boom, 1987); and one of the most important in central Amazonia (Prance et al., 1976; Mori and Lepsch-Cunha, 1995; De Oliveira and Mori, 1999) and the Chocó of Colombia (Gentry, 1982). Although most common in lowland, non-flooded forests, the family is also found in reduced numbers in cloud forests, periodically flooded riverine habitats, and in savannas. Although occasionally found in disturbed areas, species of Lecythidaceae are usually not a dominant component of secondary vegetation.
Members of New World Lecythidaceae are diversified in fruit and seed morphology and are well adapted for dispersal by both abiotic (water, wind, and gravity) and biotic (fish and mammal) agents (Prance and Mori, 1978, 1979; Mori and Prance, 1990b; Mori, 2000). Fruit and seed morphology in relation to seed dispersal has been extensively studied in general (Pijl, 1982; Murray, 1986), and the role of the seed coat in protecting the embryo and as a means of controlling dormancy and germination has long been recognized (Bewley and Black, 1994). In contrast, seed
*Corresponding author. Tel: (718) 817-8629; Fax: (718) 817-8648; E-mail: smori@nybg.org