Bot. Bull. Acad. Sin. (2002) 43: 57-62
Park and Backlund Origin of the cyathium-bearing Euphorbieae
Origin of the cyathium-bearing Euphorbieae (Euphorbiaceae): phylogenetic study based on morphological characters
Ki-Ryong Park1,* and Anders Backlund2
1Department of Biology, Kyung-Nam University, Masan 631-701, Korea
2Division of Pharmacognosy, Department of Pharmacy, Uppsala University, BMC-Biomedical center, S-751 23 Uppsala, Sweden
(Received October 6, 2000; Accepted August 24, 2001)
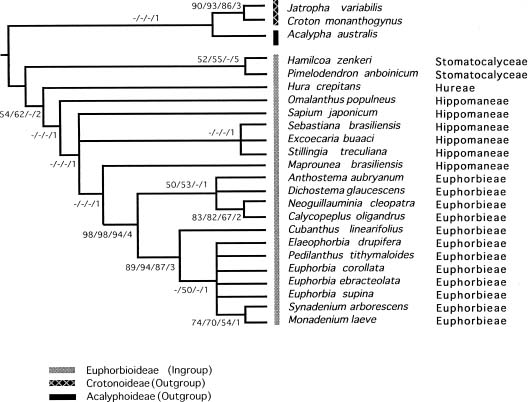
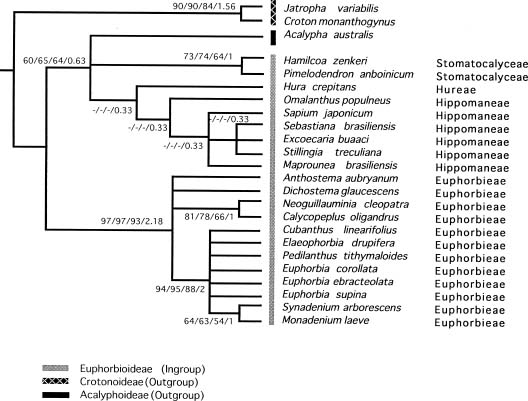
Abstract. A cladistic analysis of the subfamily Euphorbioideae was undertaken to elucidate the origin of the cyathium-bearing Euphorbieae and to provide hypotheses about evolutionary relationships within the subfamily. Twenty-one species representing most of the genera within the study group and three outgroup taxa from the subfamilies Acalyphoideae and Crotonoideae were selected for parsimony analysis. An unweighted parsimony analysis of 24 morphological characters resulted in five equally parsimonious trees with consistency indices of 0.67 and tree lengths of 39 steps. The strict consensus tree supported monophyly of the cyathium-bearing Euphorbieae. The sister group relationships of cyathium bearing Euphorbieae with Maprounea (subtribe Hippomaninae) were supported weakly, and the origin of cyathium is possibly in Hippomaneae, or in the common ancestor of Euphorbieae and remaining taxa of Euphorbioideae plus Acalyphoideae. Within the tribe Euphorbieae, both subtribes Euphorbiinae and Neoguilauminiinae are monophyletic, but the African endemic subtribe Anthosteminae is unresolved. The resulting trees support the monophyly of the tribe Stomatocalyceae while the tribe Hippomaneae does not consistently form a clade.
Keywords: Cyathium; Euphorbieae; Phylogeny.
Introduction
In a recent classification of subfamily Euphorbioideae Boiss., Webster (1975, 1994b) recognized six tribes: Stomatocalyceae (Muell. Arg.) Webster, Hippomaneae A. Juss. ex Spach, Pachystromateae (Pax & Hoffm.) Pax, Hureae Dumortier and Euphorbieae Blume. Among these the cyathium-bearing taxa are restricted to the tribe Euphorbieae. Compared to Webster's system, Pax (1924) recognized only two closely related tribes: Hippomaneae and Euphorbieae, and one remotely related tribe, Gelonieae (Muell. Arg.) Pax, on his phyletic tree. However, Gelonieae, corresponding in part to Webster's tribe Stomatocalyceae, was indicated as closely related to tribe Manihoteae (Muell. Arg.) Pax of the subfamily Crotonoideae Pax in Pax's system.
The highly specialized inflorescencecyathiumof Euphorbieae, is characterized by a central female flower and four or five lateral groups of male flowers within the involucre. Although the unique cyathium in Euphorbieae has been considered evidence of a natural group, the origin and nature of this character have been interpreted diversely (Jussieu, 1824; Mueller, 1866; Haber, 1925; Croizat, 1937, 1942; Gilbert, 1994).
Croizat (1937, 1938) pointed out a strong resemblance between Euphorbieae and Hippomaneae except with regard
to the position of a female flower. Accordingly, the Euphorbia-like cyathium results from the alteration of floral axis and the condensation of the axis of male flower in Hippomaneae. This interpretation is also supported by Pax (1924), whose idea of relationships in Euphorbiaceae indicates that Hippomaneae is most closely related to Euphorbieae.
A recent hypothesis, however, does not support the traditional relationships (Pax, 1924; Croizat, 1937, 1938) but rather suggests origin of the cyathium-bearing Euphorbieae from Crotonoideae (Gilbert, 1994). Within the Crotonoideae, Jatropha-like species with a primary female flower and lateral male flowers are considered ancestors of Euphorbieae (Gilbert, 1994). Croizat (1942) and Hurusawa (1954) on the other hand insisted that the cyathium of Euphorbieae should be derived from the pseudanthium of Dalechampia Plumier ex L. in Acalyphoideae Ascherson via Pedilanthus Necker ex Poiteau, but several authors disputed the Dalechampia origin of Euphorbieae (Venkata Rao, 1971; Webster and Webster, 1972; Webster, 1994a).
Pollen morphology indicates that Euphorbia is similar to Stillingia Garden ex L. and Sapium P. Browne of Hippomaneae in having an even margo and a similar sculpturing pattern, but its pollen clearly differs from that of Crotonoideae and Acalypoideae (e.g. Dalechampia) (Webster and Webster, 1972; Webster and Rupert, 1973; Park, 1997). The pollen grains of Crotonoideae are mostly inaperturate with triangular supratectal elements completely absent in Euphorbieae (Park and Lee, 1988; Nowicke, 1994; Webster, 1994b).
*Corresponding author. Tel: 82-55-249-2240; E-mail: park@kyungnam.ac.kr