Bot. Bull. Acad. Sin. (2002) 43: 83-91
Ho et al. Metagentiana, a new genus of Gentianaceae
Metagentiana, a new genus of Gentianaceae1
Ting Nong Ho*, Shi Long Chen, and Shang Wu Liu
Northwest Plateau Institute of Biology, Chinese Academy of Sciences, Xining Qinghai, 810001, P.R. China
(Received June 1, 2000; Accepted April 16, 2001)
Abstract. The evidence from gross morphology, floral anatomy, chromosomes, palynology, and embryology all indicates that sect. Stenogyne is discordant within the genus Gentiana and is as distinct from the other sections of Gentiana as are other genera, such as Tripterospermum and Crawfurdia. In light of these characters, sect. Stenogyne is removed from Gentiana and given generic rank as the new genus Metagentiana. It is more related to Tripterospermum and Crawfurdia than to Gentiana, though it is more primitive than the first two genera. Together with Tripterospermum and Crawfurdia the new genus forms a monophyletic group, which is the sister group to the genus Gentiana. Fourteen new combinations required at specific rank are proposed.
Keywords: Gentiana; Gentianaceae; Metagentiana; New Genus; Taxonomy.
Introduction
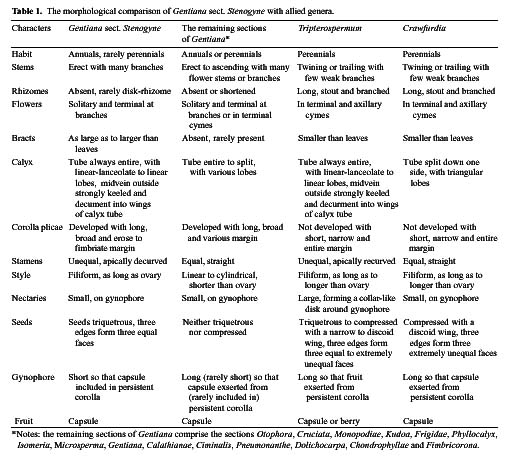
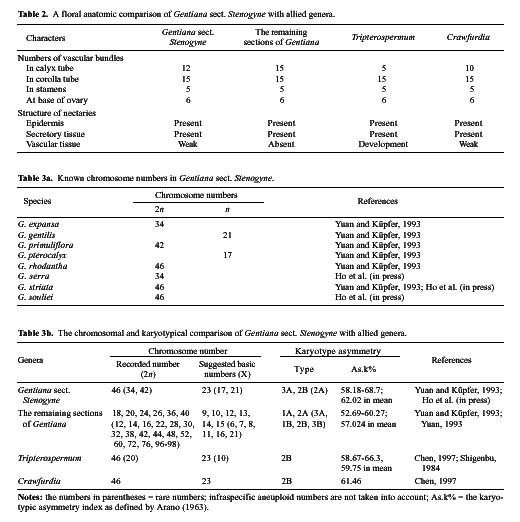
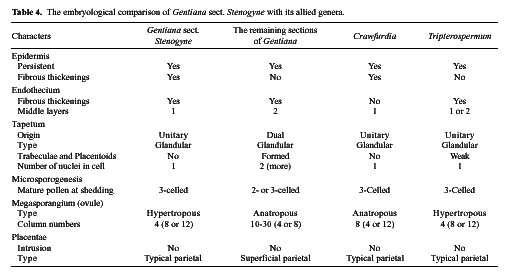
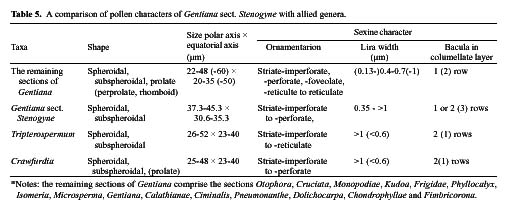
Gentiana sect. Stenogyne was established by Franchet (1884) and was revised by Kusnezov (1894). It contains fourteen species: Ten of these are highly restricted to southwestern China; two are relatively widespread in northwestern and central China; one extends to eastern Burma from southwestern China, and one is endemic to Thailand. Stenogyne is the most disputable and poorly known of Gentiana's 16 sections. Smith (1965) considered it a somewhat mysterious group in Gentiana from its morphological characters, which suggested a closer affinity with Tripterospermum or Crawfurdia than with other sections of Gentiana. Löve and Löve (1976) recommended the transfer of this section to the genus Tripterospermum, tentatively as a subgenus (Tripterospermum subgen. Stenogyne). However, they had no karyological information to make a further delimitation. Yuan and Küpfer (1993) reported chromosome numbers for six species of this section for the first time and suggested a specialized and isolated position of this section in the genus Gentiana because of its higher and obviously secondary basic chromosome numbers x=17, 21, and 23. Three years later (Yuan et al.,1996), based on the nucleotide sequence of the internal transcribed spacer (ITS) of nuclear Ribosomal DNA, they further suggested excluding sect. Stenogyne from Gentiana and classifying it as a distinct genus. However, the necessary taxonomic changes were never made. Halda (1995) made sect. Stenogyne a subgenus, still positioned in the genus Gentiana. Since 1990, we have examined gross
morphology, floral anatomy, karyology, pollen morphology and embryology for most genera in the subtribe Gentianinae and have published a series of papers (Chen et al., 1997, 1998, 1999a, 1999b, 2000; Ho and Liu, 1990, 1999; Ho and Pringle, 1995; Ho et al., 1994, 1996, 1997, 1999, 2000; Liu and Ho, 1996a, 1996b, 1997; Xue et al., 1999a, 1999b). The present paper attempts to bring together these interactive studies of various materials relating to Gentiana sect. Stenogyne with the hope of better understanding its classification and systematic position.
Character Analyses
Based on detailed comparisons of characters such as gross morphology, floral anatomy, chromosomes, pollen morphology, and embryology, this study presents a complete documentation of Gentiana sect. Stenogyne. The concise descriptions of these characters used for the present study are as follows.
Gross Morphology
Most species in Gentiana sect. Stenogyne are annual herbs, but a few are perennials with extremely short rhizomes. The stem is erect and branched. Leaves are sessile, broadly ovate to ovate-triangular. Flowers are many, with each solitary and sessile at the end of stems and branches, subtended by a pair of leaf-like bracts. The calyx is tubular; midveins of the lobes are keeled and winged into the calyx-tube. The corolla is funnelform, and its plicae is long and wide. The margin of the plicae may be erose, denticulate, or fimbriate. The stamens are unequal and more or less curved downwards unilaterally. There are 5 nectaries at the base of the ovary. The style is filiform and about as long as the ovary. The capsules are cylindrical. The seeds are triquetrous with the three edges winged or rarely unwinged. Sect. Stenogyne is very
1This project was financially supported by the Chinese National Natural Foundation.
*Corresponding author. E-mail: hotingnong@163.net