Bot. Bull. Acad. Sin. (2002) 43: 99-105
Tseng et al. SAMPL genotyping of sweet potato cultivars
Genotyping and assessment of genetic relationships in elite polycross breeding cultivars of sweet potato in Taiwan based on SAMPL polymorphisms
Yu-Tien Tseng 1, Hsiao-Feng Lo2, and Shih-Ying Hwang1,*
1Graduate Institute of Biotechnology, Chinese Culture University, 55 Hwagan Rd., Yangmingshan, Taipei, Taiwan
2Department of Horticulture, Chinese Culture University, 55, Hwagan Rd., Yangmingshan, Taipei, Taiwan
(Received September 11, 2001; Accepted November 28, 2001)
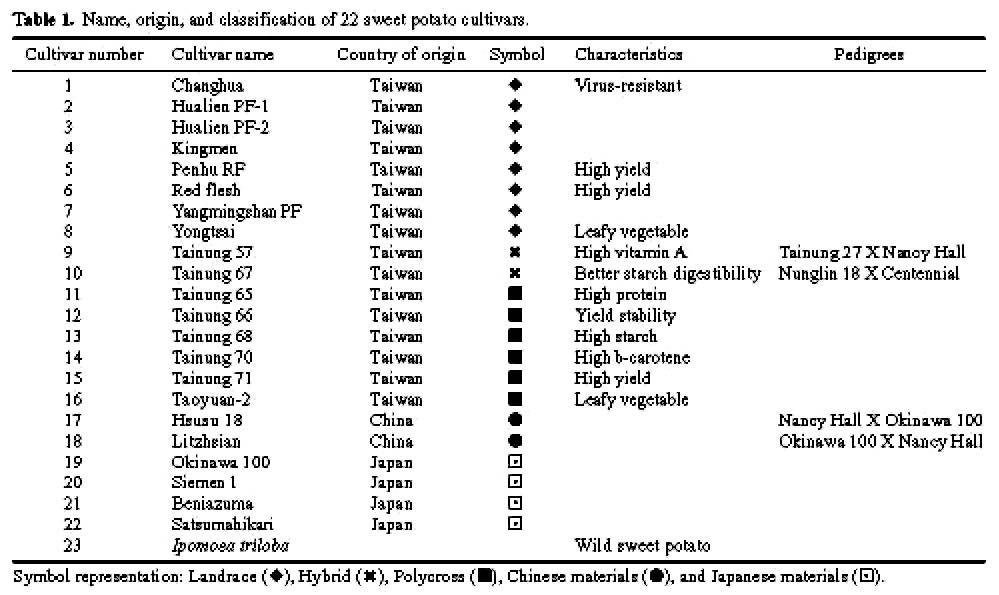
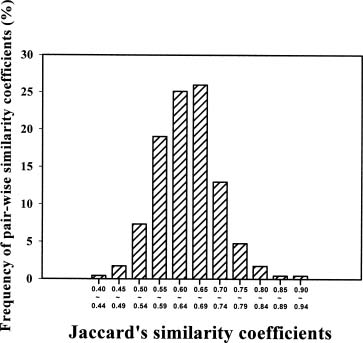
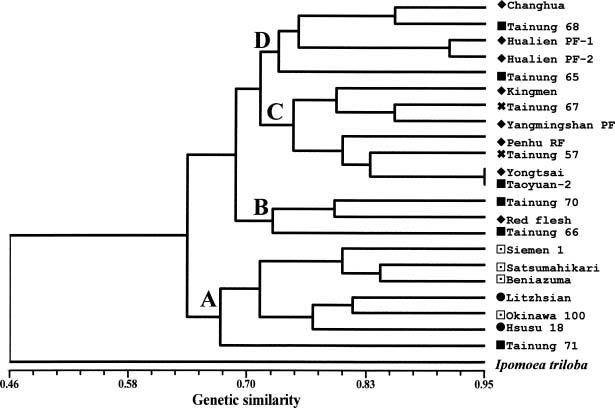
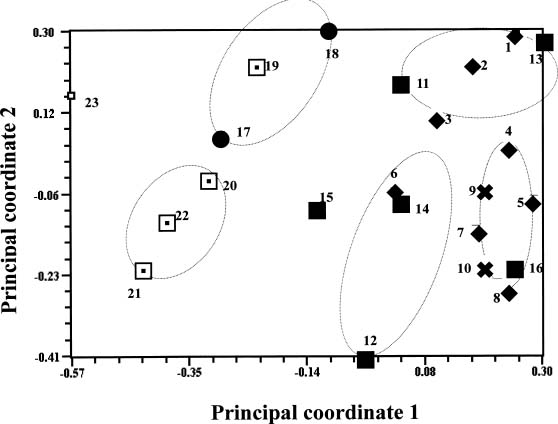
Abstract. Selective amplification of microsatellite polymorphic loci (SAMPL) markers was used to analyze the genetic relationships between 22 elite cultivars of sweet potato [Ipomoea batatas (L.) Lam.] used in polycross breeding in Taiwan. These elite cultivars included varieties introduced from China and Japan and landraces from Taiwan as well as cultivars derived either from hybrid or polycross breeding programs. Among 12 SAMPL primer pairs tested, 7 amplified 19 loci while the other 5 SAMPL primer pairs gave ambiguous banding profiles. Fifty-five alleles were amplified in total. The unweighted pair group method with arithmetic mean (UPGMA) and principal coordinate analysis (PCA) of SAMPL data suggest that Taiwan landraces are distantly related to Chinese and Japanese cultivars after many years of independent selection and may have originated from Java and Brahman. Polycross derived cultivars are closely related to different landraces. Employment of SAMPL markers is efficient compared to other molecular marker methods, such as random amplified polymorphic DNA (RAPD) and simple sequence repeat (SSR), in assessing the genetic relationships of sweet potato cultivars used in the polycross breeding program in Taiwan.
Keywords: Genetic relationships; Polycross; SAMPL; Sweet potato.
Introduction
Information on germplasm diversity and genetic relationships among elite breeding lines is critical in crop improvement. Sweet potato (2n=6X=90) (Ozias-Akins and Jarret, 1994) is the seventh most-important crop in the world and a major source of food and nutrition in developing countries (International Potato Center, 1996). Sweet potato in Taiwan was first introduced from Java and Brahman as early as the 17th century during Dutch colonization. Afterwards, the Japanese introduced sweet potato materials from Japan, the USA, China, and also from Java and Brahman. Other sweet potatoes were introduced after World War II mainly from China, Japan, and the USA. The breeding of sweet potatoes is mostly through hybrid or polycross breeding. Pre-screening of cross-compatibility is necessary, but time consuming, for the success of hybrid breeding. Jones (1965) proposed a polycross breeding scheme in which 20-30 parental lines are planted in an isolated plot and subjected to open pollination by insects and mass selection to eliminate the pre-screening of cross-compatibility. The polycross strategy has been used in Taiwan to generate many elite sweet potato cultivars such
as high yield, high b-carotene, and high protein cultivars. Despite common failure in positive identification of parental lines using morphological markers that are subject to change due to environmental factors and cultural practices (Bernatzky and Tanksley, 1989), polycross breeding has been the major breeding program practiced in Taiwan since 1966. Since then, sweet potatoes of China and Japan and landraces in addition to hybrid- and polycross-derived cultivars have dominated the polycross breeding parental lines in Taiwan.
In sweet potato, random amplified polymorphic DNA (RAPD) has been used in cultivar fingerprinting (Connolly et al., 1994; Hwang et al., 1999), estimation of genetic diversity (Sagredo et al., 1998; Zhang et al., 1998), genome mapping (Ukoskit and Thompson, 1997), and locating a root-knot nematode resistance gene (Ukoskit et al., 1997). Although open pollination by insects and mass selection are practiced in polycross breeding programs, the breeding efficiency is low. In recent years it has become well recognized that PCR-based DNA markers make powerful tools for genetic analysis and for breeding programs because of their simplicity and ease of handling. Primers designed to flank simple sequence repeats (SSRs) loci were developed (Jarret and Bowen, 1994) and used to genotype sweet potato cultivars (Hwang et al., 2002). However, Buteler et al. (1999) reported that in sweet potato, inser
*Corresponding author. Tel: (02) 28610511 ext. 628; Fax: (02) 28618266; E-mail: hsy9347@ms34.hinet.net