Bot. Bull. Acad. Sin. (2002) 43: 115-122
Hsiao et al. Bound form of plant invertase in rice
A novel bound form of plant invertase in rice suspension cells
Cheng-Chung Hsiao1, Ru-Huei Fu1, and Hsien-Yi Sung*
Biochemistry Laboratory, Department of Agricultural Chemistry, National Taiwan University, 1 Roosevelt Road Section 4, Taipei, Taiwan 106, Republic of China
(Received August 14, 2001; Accepted December 20, 2001)
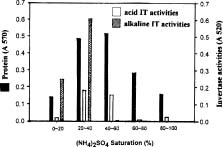
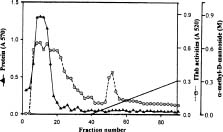
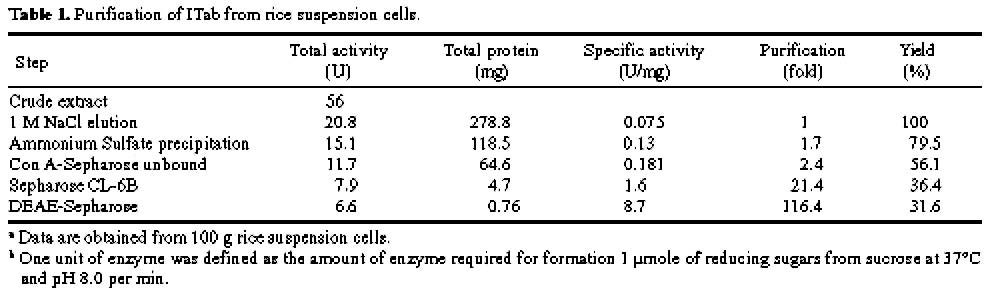
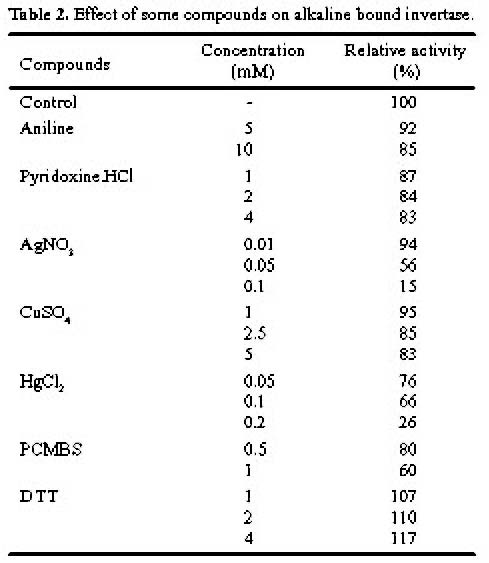
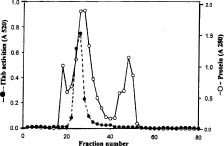
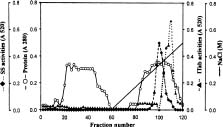
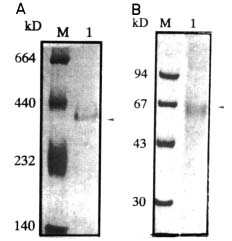
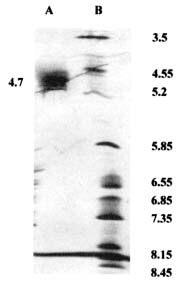
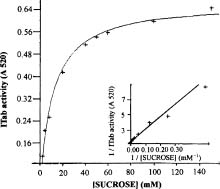
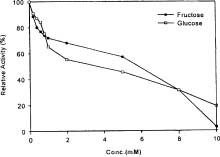
Abstract. A novel alkaline bound invertase (ITab) was purified 116 fold from crude extract of rice suspension cells (Oryza sativa L. cv. Tainong 67). The enzyme can be released by treating the cell wall and membrane fragments with 1.0 M NaCl and purified to homogeneity through the steps of ammonium sulfate precipitation, Con A-Sepharose affinity (non-retained), Sepharose CL-6B, and DEAE-Sepharose chromatographies. The FPLC and SDS-PAGE analyses revealed ITab to have a native molecular mass of about 250 kDa, and it may therefore be homotetramer composed of subunit with a mass of 60 kDa. Isoelectric focusing analysis indicated that the pI value was 4.7. The optimum pH was 7.6 and the optimum temperature was 40-50°C. ITab can hydrolyze sucrose and raffinose, but not maltose. The enzyme did not have affinity toward Con A. So it is not a glycoprotein. The Km for sucrose was 15.28 mM. Fructose and glucose were inhibitors. The enzyme was activated by additional protein such as bovine serum albumin. Tris, metal ions, and thiol reagents inhibited the enzyme activity, but activity could be restored with DTT. It is suggested that sulfhydryl-group(s) exist either in the active site or nearly active site.
Keywords: Bound form invertase; Rice suspension cells; Sucrose metabolism.
Abbreviations: Con A, concanavalin A; DTT, 1,4-dithiothreitol; ITab, alkaline bound invertase; PCMBS, p-chloromercuribenzenesulphonic acid.
Introduction
Sucrose is one of the predominant initial products of photosynthesis and serves as the major carbohydrate translocator, storage component, osmoticum, and regulator of gene expression (Koch, 1996) in higher plants. Utilization of the most sucrose depends on its cleavage into glucose and fructose, and in plants it is either invertase, also called b-D-fructofuranoside fructohydrolase (EC 3.2.1.26), or sucrose synthase (EC 2.4.1.13) that catalyzes this reaction.
Invertase isozymes can be distinguished by their subcellular localization, solubility, and optimum pH. The acid bound invertase (insoluble) with a basic pI is ionically or covalent bound to the cell wall. The acid unbound invertase (soluble) with an acidic pI is localized in the vacuole; and the alkaline unbound invertase (soluble) is thought to be cytoplasm (Tymowska-Lalanne and Kreis, 1998). Acid invertases are glycosylated forms and also hydrolyze other b-fructofuranoside-containing oligosaccharides such as raffinose and stachyose, while alkaline invertases are most likely a non-glycosylated form and appear to be sucrose specific (Sturm, 1999).
Acid invertases are synthesized as preproproteins, with long leader sequences, which are cleaved off during transport and protein maturation. The amino acid sequences of the plant acid invertases share a few highly conserved motifs: b-fructosidase motif (NDPNG/A), close to the N terminus of the mature proteins, and cysteine catalytic site (MWECV/PDF), located closer to the middle of mature proteins (Sturm, 1999). It is pertinent that almost all cell wall bound invertases have a proline residue in the cysteine catalytic site while vacuolar invertases have a valine residue (Tymowska-Lalanne and Kreis, 1998). In addition, most plant invertases have a third domain, SI/VVESF, but its function is not clear.
In connection with the various roles sucrose plays in plants, invertases may have several different functions (Sturm and Tang, 1999). The vacuolar invertases not only mobilize sucrose and control sugar composition in fruits and storage organs, but also play a role in osmoregulation, cell enlargement, and cold sweetening. Cell wall bound invertases are considered to be key enzymes involved in sucrose unloading, wounding, infection response, and cell differentiation. The cytosolic invertase is most likely involved in the regulation of intracellular glucose and fructose levels in mature tissues and channeling sucrose into catabolism, in which sucrose synthase activity is low. The presence of multiple isoforms of invertase might confer a great flexibility to the control of sugar metabolism,
1Both authors contributed equally to this work.
*Corresponding author. Tel: 886-2-23634729; Fax: 886-2-23634729; E-mail: SUNGHY@ccms.ntu.edu.tw