Bot. Bull. Acad. Sin. (2002) 43: 161-170
Chang et al. Taxonomy of Codium geppiorum complex
A taxonomic and karyological study of the Codium geppiorum complex (Chlorophyta) in southern Taiwan, including the description of Codium nanwanense sp. nov.
Jui-Sheng Chang1, Chang-Feng Dai1, *, and Jeng Chang2
1Institute of Oceanography, National Taiwan University, Taipei 106, Taiwan
2Institute of Marine Biology, National Taiwan Ocean University, Keelung 202, Taiwan
(Received Feburary 13, 2001; Accepted September 26, 2001)
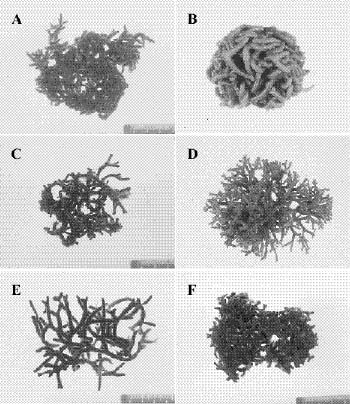
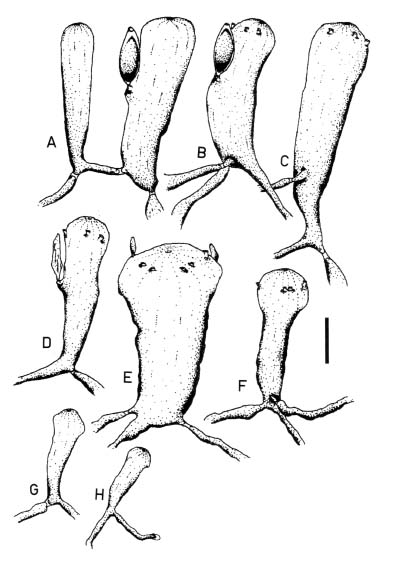
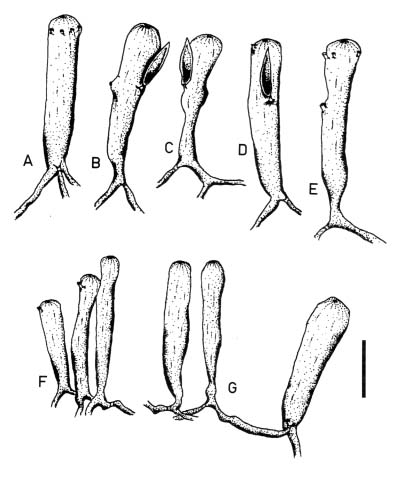
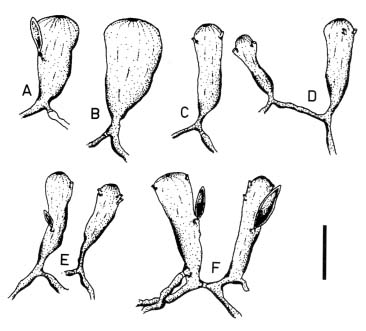
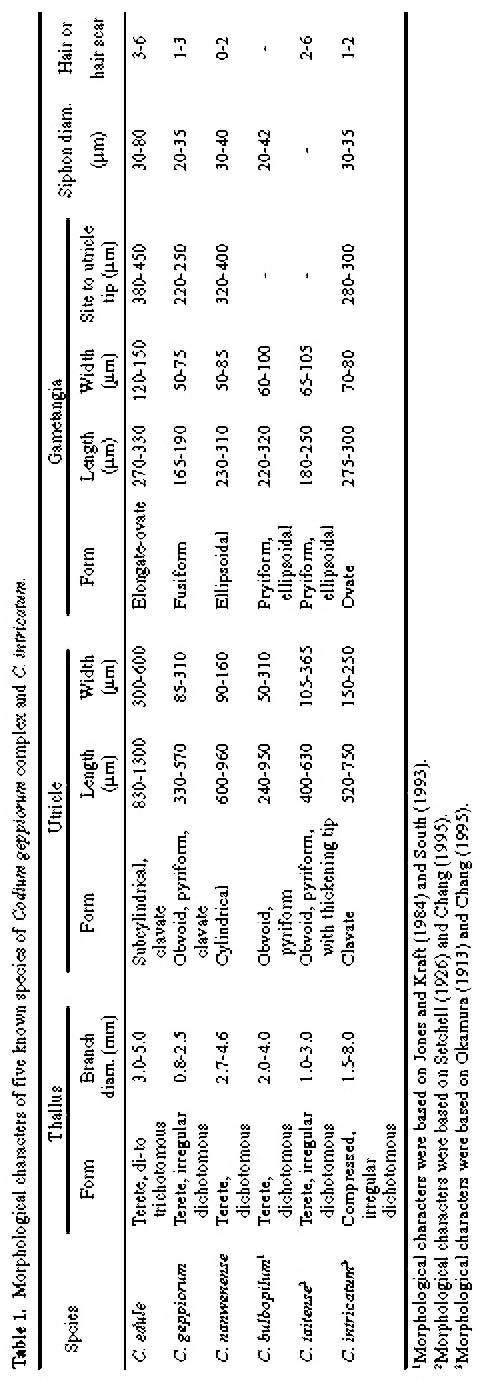
Abstract. Morphological variations of the Codium geppiorum complex from southern Taiwan were examined, and their karyotypes were studied by microspectrophotometry. Morphological analysis distinguished three prostrate Codium species, including C. edule P.C. Silva, C. geppiorum O.C. Schmidt, and a new taxon, C. nanwanense. Although these species show an anatomical continuum as the length of their utricles increases with the thickness of the branches, there are some apparent differences of thallus type and utricle form. The utricles of C. edule are about 1000 µm or more in length, and with abundant hair scars on the utricle tip. Codium geppiorum has a typical pyriform utricle, which is often less than 500 µm in length. The utricles of C. nanwanense are cylindrical and medium-sized, between C. edule and C. geppiorum. Karyological study on the three Codium species also showed an obvious divergence in nuclear size and relative fluorescent unit (RFU). Codium edule has the largest nucleus (8.90 µm in average length) and expectedly the highest RFU among the three species. The ratio of nuclear long axis vs. width is about 3.01, 1.46 and 1.00 for C. edule, C. geppiorum, and C. nanwanense. This paper summarizes the taxonomic characters of these species and provides microfluorescence data useful in distinguishing members of the Codium geppiorum complex.
Keywords: Codium geppiorum complex; Codium nanwanense sp. nov.; Karyology; Microfluorescence; Morphology.
Introduction
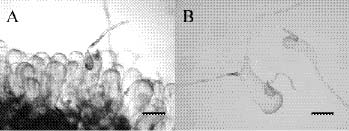
The coenocytic, siphonous green alga Codium Stackhouse (Bryopsidales, Codiaceae) is one of the most common and widespread seaweeds in the world (Silva, 1951, 1952). There are about 100 described species, which frequently grow on rocky shores in tropical and temperate areas (Silva, 1962, 1992). Codium is characterized by its spongy thallus, composed of a colorless medulla of densely intertwined filaments and a green palisade-like cortex composed of vesicles called utricles. Thallus type varies from flattened crusts to globular and decumbent or erect systems, consisting of cylindrical branches or unbranched laminate blades (Bold and Wynn, 1985). The coenocytic utricles have discoid chloroplasts devoid of pyrenoids. Dark-green female and yellowish-green male gametangia are commonly borne on the distal half of the utricles. Codium exhibits marked anisogamy and possesses biflagellate gametes (Silva, 1952, 1982; Van den Hoek et al., 1995; Lee, 1999).
The taxonomy of Codium is complicated by the high degree of anatomical and habit variability within and be
tween populations (Silva, 1951). Most of the Codium species are distinguished on the basis of gross morphological features combined with anatomical details, such as size and form of utricles and reproductive organs, structure of the apex of the utricles, position of utricle hairs and septa, and ontogenetic patterns (Silva, 1982). Morphological variations of thallus type and utricle form present taxonomic difficulties for some closely related Codium species because an anatomical continuum appears as the length of their utricles increases with thickness of the branches. Codium fragile (Suringar) Hariot with several subspecies is one such group within this diverse genus and is an interesting taxonomic case study because of its widespread distribution and invasive characteristics (Trowbridge, 1998). Other groups, such as the C. arabicum complex and the C. geppiorum complex, are still poorly delineated and often confused with each other due to the utricle size continuum in similar morphological characters. Although some molecular studies have been performed, they remain focused on the species C. fragile and its subspecies (Manhart et al., 1989; Goff et al., 1992).
Codium geppiorum O.C. Schmidt is a substitute name for C. divaricatum A. Gepp & E.S. Gepp, which is a later homonym of C. divaricatum (C. Agardh) Biasoletto (Schmidt, 1923). Codium geppiorum is a complex of repent, procumbent, or decumbent anastomosing types
*Corresponding author. Tel: 02-23916693; Fax: 02-23916693; E-mail: corallab@ccms.ntu.edu.tw