Bot. Bull. Acad. Sin. (2002) 43: 171-180
Wu et al. Co-dominant RAPDs of rice morphological genes
Co-dominant RAPD markers closely linked with two morphological genes in rice (Oryza sativa L.)
Ju-Yu Wu, Hsin-Kan Wu, and Mei-Chu Chung*
Institute of Botany, Academia Sinica, Nankang, 11529, Taipei, Taiwan, Republic of China
(Received October 9, 2001; Accepted March 1, 2002)
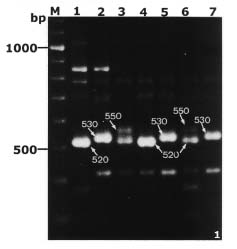
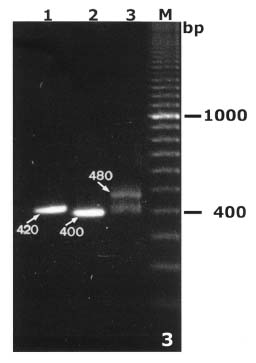
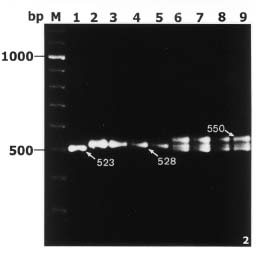
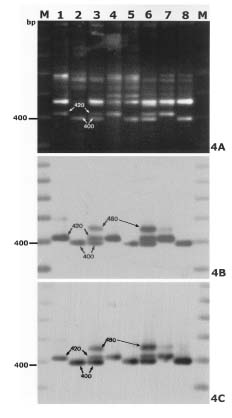
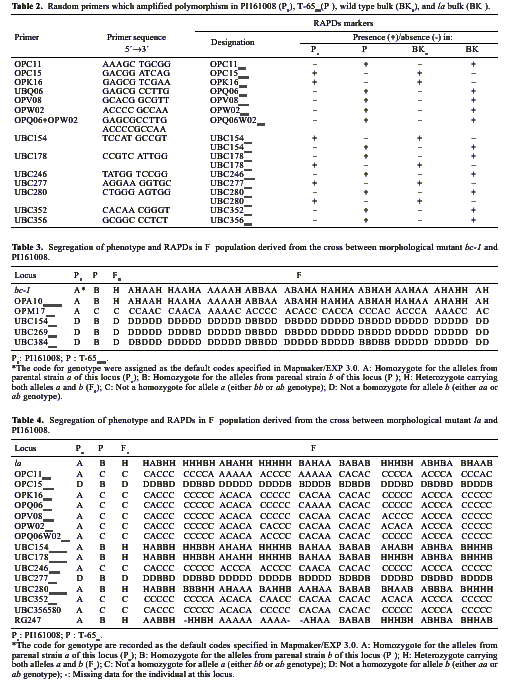
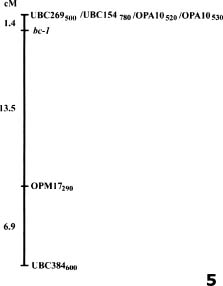
Abstract. By bulked segregant analysis, we screened for markers linked to two morphological genes, brittle culm (bc-1) and lazy (la). We identified four dominant and two co-dominant RAPD (randomly amplified polymorphic DNA) markers for bc-1 and eleven dominant and six co-dominant RAPD markers for la. One of the co-dominant RAPD markers, located 1.4cM from bc-1, was converted into co-dominant SCAR (sequence characterized amplified region). The six co-dominant markers were linked to la at various distances; two of which spanned a region with many RAPD markers. These co-dominant RAPD markers were detected by size and were a result of sequence modification between, not within, the priming sites. Non-parental bands were observed when the F1 and the heterozygous F2 RAPD and SCAR products were analyzed electrophorectically. Possible reasons for these non-parental bands were examined by sequence analysis, DNA annealing experiments, and Southern hybridization. The results indicated they were heteroduplex DNA formed by identical fragments amplified from allelic sequences. The presence of non-parental bands is further evidence of the co-dominance of the markers of interest. The co-dominant RAPD markers developed in this study overcome many of the limitations of RAPDs, and are readily verifiable and applicable. With these and additional PCR based markers, gene mapping or marker aided selection can be made more efficient.
Keywords: Co-dominant RAPD markers; Linkage; Morphological mutant; Oryza sativa; SCAR.
Introduction
Rice (Oryza sativa L.), with its small diploid genome, consists of 12 well defined genetic linkage groups (Bennett and Smith, 1976; Kinoshita, 1986; Khush and Singh, 1986; Wu et al., 1988; Ranjhan et al., 1988; Arumuganthan and Earle, 1991). A classical linkage map updated by the Rice Genetics Cooperative carries 209 genes, which include morphological, disease resistance and isozyme markers (Kinoshita, 1998). A molecular linkage map, based on restriction fragment length polymorphism (RFLP), was first reported by McCouch et al. (1988). Subsequently, several molecular genetic maps have been developed by Saito et al. (1991), Kurata et al. (1994), Causse et al. (1994), and Harushima et al. (1998). While the number of markers that can be assigned to these linkage maps is increasing (URL for http://shigen.lab.nig.ac.jp; http://www.gramene.org), the markers are not readily cross-referenced. In order to more fully exploit the information so accumulated, efforts have been made to integrate the classical linkage and molecular maps (Kishimoto et al., 1993; Abenes et al., 1994; Yu et al., 1995; Yoshimura et al., 1997). Cho et al. (1998) have integrated microsatellite, AFLP (amplified fragment length polymorphism) and SSLP (simple-sequence length polymorphism) markers into an RFLP map of rice with the aim of filling gaps, enhancing the level of saturation, and providing a reasonable start
ing point for high resolution QTL analysis, gene isolation, and molecular breeding.
RAPD (randomly amplified polymorphic DNA, Williams et al., 1990; Welsh and McClelland, 1990) markers are also promising for this purpose. RAPD requires minute quantities of DNA for detection of polymorphism based on the presence (dominant) or absence (recessive) of particular bands in electrophoresis. However, its limitation is that being dominant, RAPD markers provide less information for evaluation in linkage analysis than co-dominant RFLP markers. The recombination between dominant RAPD markers and target genes cannot be estimated effectively in a segregating population, and multi-loci inheritance may restrict the usefulness of RAPD markers. In the latter case, PCR (polymerase chain reaction) analysis based on SCAR (sequence characterized amplified region, Paran and Michelmore, 1993) would be necessary (Nair et al., 1995; Nair et al., 1996). RAPD analysis combined with BSA (bulked segregant analysis) has also been used to focus on regions of interest or areas sparsely populated with markers (Giovannoni et al., 1991; Michelmore et al., 1991; Zhang et al., 1994; Zhang et al., 1996). BSA provides information simultaneously on polymorphism of the parents, and possible linkage between a marker and a targeted gene; only two bulks and the parents are needed, reducing the cost and labor requirement several fold compared to conventional methods. In rice, RAPD markers closely linked to genes resistant to gall midge (Nair et al., 1995; Nair et al., 1996), to bacterial blight (Zhang et al., 1996), and to brown planthopper (Jena et al., 1998) have been successfully screened by these approaches.
*Corresponding author. Tel: 886-2-27899590 ext. 114; Fax: 886-2-27827954; E-mail: bomchung@ccvax.sinica.edu.tw