Bot. Bull. Acad. Sin. (2002) 43: 181-185
Yang et al. Photoprotective mechanism of exogenous b-carotene
Effects of b-carotene feeding on chlorophyll fluorescence, zeaxanthin content, and D1 protein turnover in rice (Oryza sativa L.) leaves exposed to high irradiance
Cheng-Wei Yang*, Chang-Lian Peng, Jun Duan, and Yi-Zhu Chen
South China Institute of Botany, The Chinese Academy of Sciences, Guangzhou 510650, The People's Republic of China
(Received October 19, 2001; Accepted March 19, 2002)
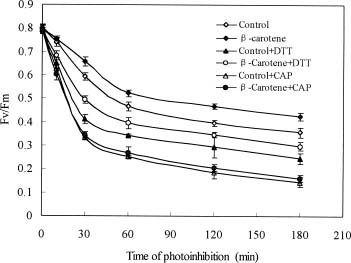
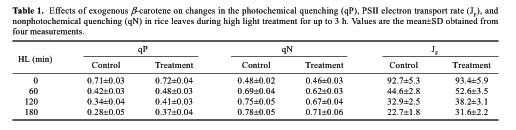
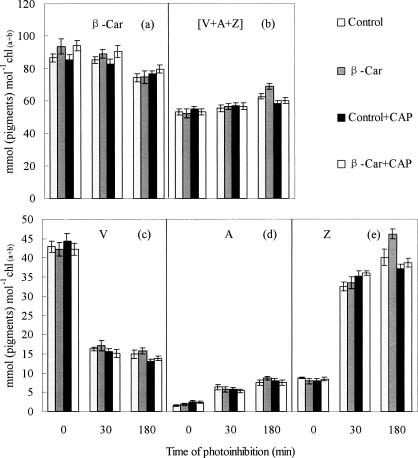
Abstract. To examine the mechanism of photoprotective effect of exogenous b-carotene against photoinhibition under strong light conditions, leaves of rice (Oryza sativa L.) were fed with 30 mmol/L b-carotene through the transpiration stream at a PPFD of 20 µmol m-2 s-1 for 3 h. The leaves were then exposed to strong light at a PPFD of 2000 µmol m-2 s-1 for another 3 h. The photo-induced decrease in activity of PS2 in b-carotene fed leaves, as estimated in terms of the changes of Fv/Fm, photochemical quenching (qP), and PSII electron transport rate (JF), was smaller than that in control leaves during exposure to strong illumination. However, the presence of chloramphenicol produced no significant difference. Furthermore, feeding of b-carotene increased endogenous b-carotene content. In addition, the content of zeaxanthin and the xanthophyll cycle pigments pool size (V+A+Z) were increased in comparison with control leaves when the leaves were exposed to high light for 3 h. However, no significant difference was observed in the presence of chloramphenicol. These results indicate that the photoprotective effect of b-carotene feeding can be partially explained by the conversion of b-carotene to zeaxanthin, a process involved in the rapid D1 protein turnover during the reassembly of PS2 in high irradiance.
Keywords: b-Carotene; Chlorophyll fluorescence; D1 protein turnover; High irradiance; Xanthophyll cycle; Zeaxanthin.
Abbreviations: A, antheraxanthin; b-Car, b-carotene; CAP, chloramphenicol; DTT, dithiothreitol; Fv/Fm, maximum photochemical efficiency of PS2; PPFD, photosynthetic photon flux density; PS2, photosystem 2; qP, Photochemical quenching; qN, non-photochemical quenching; JF, PSII electron transport rate; V, violaxanthin; Z, zeaxanthin.
Introduction
The inhibition of photosynthetic activity by high irradiance is a long-known phenomenon (Kandler and Sironval, 1959). Plants have developed a wide range of mechanisms that ameliorate photoinactivation by converting excitation energy harmlessly into heat, thereby preventing the formation of reactive oxygen species and protecting PS2 against photoinactivation during high light stress (for an overview see Niyogi, 1999).
As a result of adaptation to environmental conditions, higher plants possess several enzymatic and non-enzymatic scavenging systems to minimize the deleterious effects of reactive oxygen species. b-Car is an important membrane-bound antioxidant in plant tissue that can quench 1O2 produced from interaction of 3P680 and O2 in the PS2 reaction center (Telfer et al., 1994). There are two b-Car molecules in the reaction center of PS2 bound to the D1 and D2 protein (Nanba and Satoh, 1987), but they are not involved in the triplet quenching of P680 (Telfer et al., 1994), and
they seem to be lost and hydroxylated to zeaxanthin during the degradation of damaged D1 protein under high irradiance (Trebst and Depka, 1997). The content of b-Car decreased, while the relative content of pigments of the xanthophyll cycle was almost stable and the extent of photoinhibition increased during the senescence of rice leaves (Yang et al., 2001). Deo and Biswall (2001) suggested that b-Car may contribute to the assembly and stability of the D1 protein during senescence and water stress in clusterbean cotyledons. As shown earlier by Markgraf and Oelmueller (1991), b-Car was obligatory in the assembly of PS2 in the greening of etiolated tissue. Recently, the role of the b-Car in the rapid turnover and assembly of the D1 protein into the PS2 center of a green alga has been studied under photoinhibitory conditions (Depka et al., 1998; Deo and Biswal, 2001).
That b-Car is destroyed under high light has been recognized for decades. The reduction of the b-Car pool (Demmig-Adams, 1990), in parallel to an increase of the xanthophyll cycle pool size was reported (Trebst and Depka, 1997). However, little information is known about the relationship among b-Car biosynthesis, zeaxanthin formation, and D1 protein turnover under high light. In the present paper, we report exogenous feeding of b-Car on the
*Corresponding author. Tel: +86-20-87705626 ext. 405; Fax: +86-20-87639415; E-mail: yangcenw@scib.ac.cn; Yangcw72@yahoo.com