Bot. Bull. Acad. Sin. (2002) 43: 211-218
Li et al. Mycoparasitism of Gliocladium roseum on Botrytis cinerea
Ultrastructural study of mycoparasitism of Gliocladium roseum on Botrytis cinerea
G.Q. Li1, H.C. Huang*, E.G. Kokko, and S.N. Acharya
Agriculture and Agri-Food Canada, Lethbridge Research Centre, P.O. Box 3000, Lethbridge, Alberta T1J 4B1, Canada
(Received September 26, 2001; Accepted January 17, 2002)
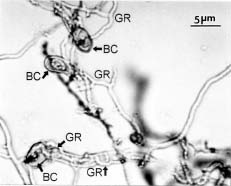
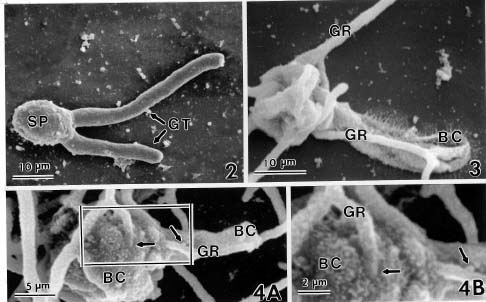
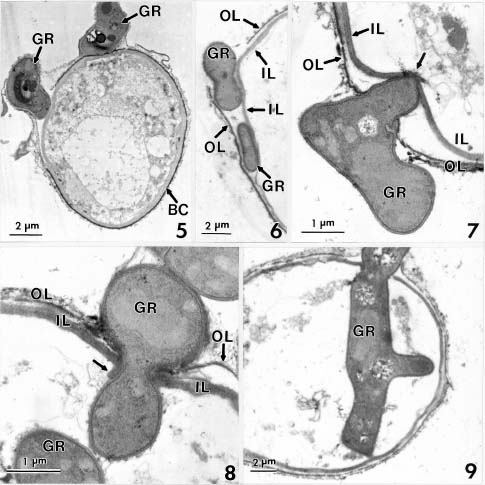
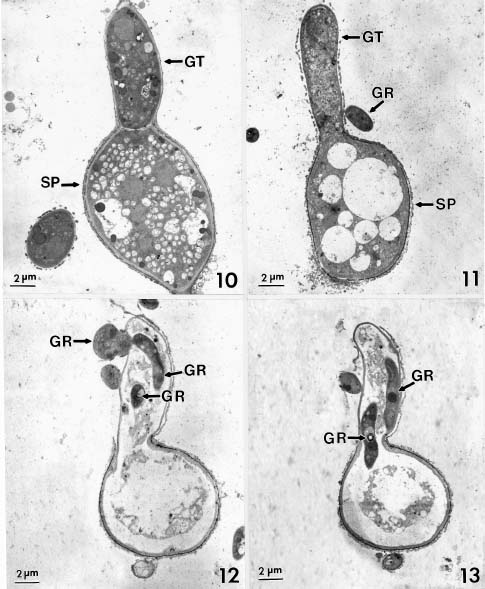
Abstract. Conidia and germ tubes of Botrytis cinerea were highly susceptible to infection by the mycoparasite Gliocladium roseum, when both organisms were grown as a mixed culture in sterile water containing 8-9×104 pollen grains of alfalfa per ml solution. Scanning electron microscopic studies revealed that infection of conidia and germ tubes occurred through direct penetration by hyphal tips of G. roseum without the formation of appressoria. Transmission electron microscopic studies indicated indentation and rupture of the host cell walls at penetration sites. The parasitized conidia and germ tubes of B. cinerea showed signs of cytoplasmic disintegration and the presence of hyphae of G. roseum.
Keywords: Alfalfa pollen; Botrytis cinerea; Gliocladium roseum; Mycoparasitism.
Introduction
Botrytis cinerea Pers.: Fr. is a worldwide ubiquitous plant pathogen, which often causes severe losses to numerous crops (Jarvis, 1977). The pathogen causes shoot blight, blossom blight, and pod rot in seed alfalfa (Medicago sativa L.) (Stuteville and Erwin, 1990). Recent surveys showed that Botrytis blossom blight can be a serious problem for alfalfa seed production in western Canada, especially in the irrigated areas (Gossen et al., 1996, 1997, 1999).
Previous reports indicated that exogenous nutrients from plant tissues, such as senescent petals and pollen grains, are required for the germination of conidia, the subsequent growth and development of germ tubes and hyphae of B. cinerea, and for the infection of host plants by this fungus (Ogawa and English, 1960; Bachelder and Orton, 1962; Chou and Preece, 1968; McClellan and Hewitt, 1973; Huang et al., 1998, 1999, 2000). Moreover, Huang et al. (1999) observed that pollen grains of alfalfa were susceptible to infection by B. cinerea. Thus, pollen may play a significant role in the epidemics of Botrytis blossom blight of various crops including alfalfa (Ogawa and English, 1960; Bachelder and Orton, 1962; McClellan and Hewitt, 1973; Huang et al., 1998, 2000).
Gliocladium roseum Bainer, now reclassified as Clonostachys rosea (Link: Fr.) Schroers, Samuels, Seifert & W. Gams (Schroers et al., 1999), is a destructive
mycoparasite against several plant pathogenic fungi including Botrytis spp. (Barnett and Lilly, 1962; Pugh and Van Eden, 1969; Richard et al., 1974; Walker and Maude, 1975; Pachenari and Dix, 1980; Turhan, 1993; Yu and Sutton, 1997). It was reported as one of the promising biocontrol agents for control of B. cinerea (Sutton et al., 1997; Köhl et al., 1998). Studies by light microscopy showed that hyphae of G. roseum can parasitize B. cinerea (Yu and Sutton, 1997) and B. allii (Walker and Maude, 1975; Pachenari and Dix, 1980). Pachenari and Dix (1980) reported that cell wall degrading enzymes such as b-(1,3)-glucanase and chitinase, and other toxic substances produced by G. roseum were associated with the infection of B. allii by G. roseum. However, no information on the infection process of B. cinerea by G. roseum is available at the ultrastructural level. The objective of this study was to investigate the infection of conidia and germ tubes of B. cinerea by G. roseum using scanning and transmission electron microscopic studies.
Materials and Methods
Strains of Fungal Cultures
Botrytis cinerea strain LRC 2421 isolated from an infected plant of lentil (Lens culinaris Medik) in Alberta, Canada, and G. roseum strain GR-8 isolated from a decayed sclerotium of Sclerotinia sclerotiorum (Lib.) de Bary in Wuhan, China, were used in this study. For production of conidia of B. cinerea, an agar plug (5 mm in diam.) containing mycelial mats was removed from a 2-day-old culture grown on potato dextrose agar (PDA, Difco Laboratories, Detroit, MI, USA) and transferred onto PDA in a Petri dish (90 mm in diam.). Ten dishes were inoculated each time. After incubation at 20°C under fluores
1Present address: Department of Plant Protection, Huazhong Agricultural University, Wuhan, 430070, P.R. China.
*Corresponding author. Tel: 403-317-2226; Fax: 403-382-3156; E-mail: huangh@em.agr.ca. LRC Contribution No.: 387-01060.