Bot. Bull. Acad. Sin. (2002) 43: 231-240
Gituru et al. Reproductive biology of Caldesia grandis
Pollination ecology, breeding system, and conservation of Caldesia grandis (Alismataceae), an endangered marsh plant in China
Wahiti Robert Gituru, Qing-Feng Wang*, Yong Wang, and You-Hao Guo
College of Life Sciences, Wuhan University, Wuhan 430072, P.R. China
(Received November 12, 2001; Accepted March 27, 2002)
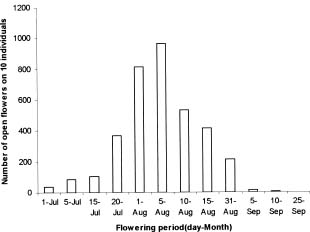
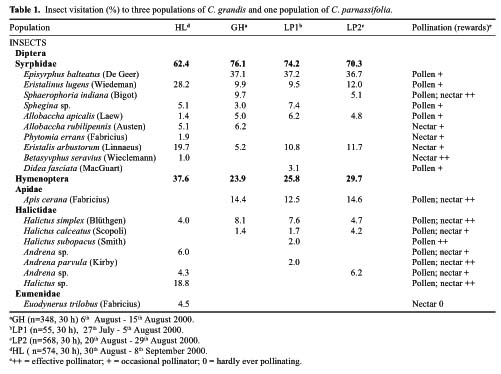
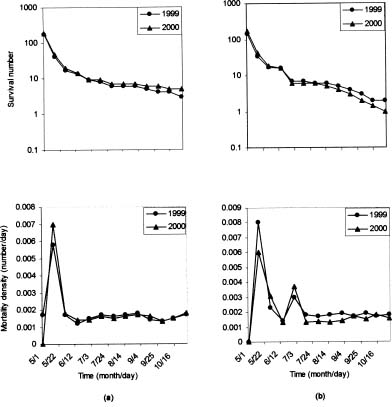
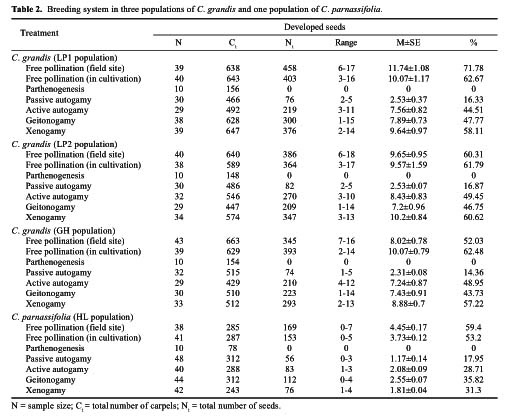
Abstract. Caldesia grandis Samuelsson is an endangered wetland herb on the brink of extinction in the vast area of Mainland China, which holds close to one eighth of the world's vascular plant species. The pollination ecology and breeding system of C. grandis were investigated from three natural populations occurring in two wetlands near the top of Mangshan Mountain in Hunan Province, central China. The species is in flower from early July to late September with a peak in August. The process of flower anthesis in C. grandis begins at about 10.00 am and lasts about four-and-one-quarter h. The flowers lasted ca. 5.5 h. Caldesia grandis is self-compatible; however, autogamy resulted in lower seed set than geitonogamy and xenogamy as well as free pollination. Both pollen viability and the seed set in open-pollinated controls at the same site were typically high (65.44% and 71.78% respectively). Mean pollen: ovule ratios in the three populations ranged from 901.75 to 931.354. No seed germination was observed, either in the field or in laboratory experiments. Propagation is achieved through turions, which commonly occur in the inflorescences. Flies (Insecta; Diptera) were the most frequent visitors to the flowers of C. grandis; however, bees (Insecta; Hymenoptera) composed a larger proportion of effective pollinators. Existence analysis revealed a peak of mortality density in mid-May corresponding to an environmental sieve in the period following the establishment of plantlets. An updated diagnostic of the conservation status of C. grandis in China was formulated. The main threats to this species are catastrophic events, especially human impact activities (agriculture, hobbyists' collections, and infrastructure construction). Establishment of a monitoring program is proposed, and both in-situ and ex-situ strategies are recommended for effective conservation of C. grandis.
Keywords: Alismataceae; Breeding system; Caldesia grandis; Conservation; Endangered; Pollination; Rare endemic.
Introduction
The genus Caldesia Parl. (Alismataceae) includes three species, all of which are restricted to the Old World. Caldesia grandis Samuelsson is confined to mountainous bogs and marshes in Southeast Asia. In comparison to the other two species in the genus, C. grandis has received less attention from researchers (Lai, 1977). This is presumably due to its limited geographical distribution (Figure 1). Furthermore, it is a mountain species, occurring at altitudes of 1,000-1,500 m in localities that are frequently not easily accessible. Caldesia grandis has been found in China and the eastern Himalayas, but not in peninsular India (Cook, 1996), Japan or Myanmar. However, only a few records of localities of the species exist. In China the species has been listed as endangered (Fu and Jing, 1992). There is a dearth of information on C. grandis in the botanical lit
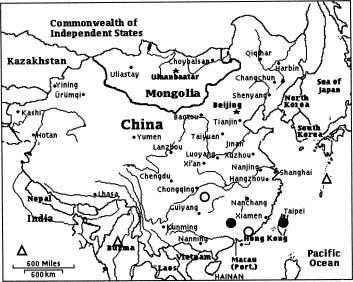
Figure 1. Global distribution of C. grandis: ~, Populations now surviving in China; , Location of recorded populations now decimated in China; r, Recorded populations outside China.
*Corresponding author. Tel: +86-27-87682869; Fax: +86-27-87669560; E-mail: wangqf97@public.wh.hb.cn