Bot. Bull. Acad. Sin. (2002) 43: 261-269
Lin et al. Construction of infectious transcripts of ZYMV
Construction of in vitro and in vivo infectious transcripts of a Taiwan strain of Zucchini yellow mosaic virus
Shih-Shun Lin1, Roger F. Hou1, and Shyi-Dong Yeh2,*
1Graduate Institute of Agricultural Biotechnology, and 2Department of Plant Pathology, National Chung Hsing University, Taichung, Taiwan 402, Republic of China
(Received September 5, 2001; Accepted May 13, 2002)
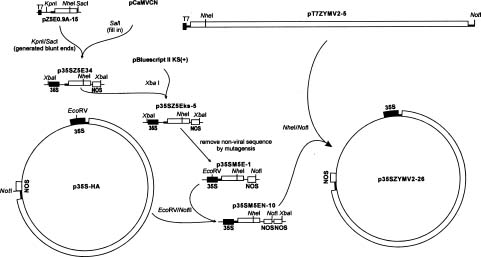
Abstract. The full-length cDNA of a Taiwan strain of Zucchini yellow mosaic virus (ZYMV TW-TN3) was constructed from five overlapping cDNA clones downstream from the bacteriophage T7 promoter in plasmid pT7ZYMV2-5. The plasmid was able to generate an in vitro transcript corresponding to TW-TN3 (9591 nt) with one extra guanosine residue at the 5´ terminus and a poly(A)95 tract at the 3´ end. In addition, pT7ZYMV2-5 was used for the construction of p35SZYMV2-26 that contained the full-length cDNA of TW-TN3 with a Cauliflower mosaic virus (CaMV) 35S promoter and a nopaline synthase (nos) terminator. The capped in vitro transcript generated from pT7ZYMV2-5 and the purified DNA of p35SZYMV2-26 were introduced into zucchini squash plants by mechanical inoculation and particle bombardment, respectively. Both in vitro and in vivo transcripts induced systemic symptoms on zucchini squash 4 to 6 days after inoculation. In addition, both transcripts also induced local lesions on plants of Chenopodium quinoa by mechanical inoculation. The results of infectivity assay, symptomatology, and serologically specific electron microscopy indicated that the in vitro and in vivo TW-TN3 transcripts derived from pT7ZYMV2-5 and p35SZYMV2-26, respectively, are infectious. The ability to generate biologically functional transcripts from the constructed cDNA clones is a significant step for molecular analyses of TW-TN3.
Keywords: In vitro; In vivo; Infectious transcripts; ZYMV TW-TN3.
Introduction
Zucchini yellow mosaic virus (ZYMV) is a member of the genus Potyvirus, the largest group among plant viruses, and is the major limiting factor for production of cucurbits worldwide. The disease is characterized by pronounced reduction in plant growth, yellow mosaic and distortion of leaves, and malformation of fruit (Lisa et al., 1981; Provvidenti et al., 1984). In Taiwan the disease caused by ZYMV has been considered one of the major limiting factors for the production of cucurbits (Lin et al., 1998; Lin et al., 2000). ZYMV particles are flexuous rods, 750 nm in length, and contain a monopartite genome consisting of a positive-sense ssRNA with a 5´ genome-linked protein (VPg) (Siaw et al., 1985; Riechmann et al., 1989; Murphy et al., 1990) and a 3´ poly (A) tract (Hari et al., 1979).
Infectious transcripts generated from cDNA clones of plant RNA viruses have been described for members of most plant virus groups (Boyer and Haenni, 1994). Among potyviruses, transcripts have been synthesized in vitro from full-length cDNA clones with bacterial phage promoters and proved to be infectious for Plum pox virus (PPV) (Riechmann et al., 1990), Zucchini yellow mosaic virus (ZYMV) (Gal-On et al., 1991), Tobacco etch virus (TEV) (Dolja et al., 1992), Pea seed-borne mosaic virus (PSbMV) (Johansen et al., 1996), Peanut stripe virus (PStV) (Flasinski
et al., 1996), Potato virus A (PVA) (Puurand et al., 1996), Tobacco vein mottling virus (TVMV) (Domier et al., 1989; Nicolas et al., 1996), Potato virus Y (PVY) (Jakab et al., 1997), Turnip mosaic virus (TuMV) (Sanchez et al., 1998) and Papaya ringspot virus (PRSV) (Chiang and Yeh, 1997). In vivo infectious transcripts, which are driven by a Cauliflower mosaic virus (CaMV) 35S promoter that can be recognized by host RNA polymerase, have also been reported for PPV (Maiss et al., 1992), ZYMV (Gal-On et al., 1995), Potato virus Y (PVY) (Fakhfakh et al., 1996), PSbMV (Johansen, 1996), Clover yellow vein virus (ClYVV) (Takahashi et al., 1997), PVY (Jakab et al., 1997), TuMV (Sanchez et al., 1998), Lettuce mosaic virus (LMV) (Yang et al., 1998), Plum pox virus (PPV) (Lopez-Moya and Garcia, 2000), TuMV (Sanchez et al., 1998) and PRSV (Chiang and Yeh, 1997). In these cases, the plasmids can be directly introduced to host plants to induce infection.
The complete nucleotide sequence of a severe Taiwan strain ZYMV TW-TN3 has previously been determined by our laboratory (Lin et al., 2001). Comparison of the coat protein (CP) gene and the P1 gene of TW-TN3 with those of other reported ZYMV isolates classified all sequenced ZYMV isolates in four genotypes, among which TW-TN3 and most of ZYMV isolates from Taiwan were placed in genotype I (Lin et al., 2000). The in vitro and in vivo infectious clones of ZYMV have been reported for an Israel isolate (IL) (Gal-On et al., 1991; Gal-On et al., 1995) that was placed in genotype II with closer relationships to US isolates (Lin et al., 2000).
*Corresponding author. Tel: 886-4-2287-7021; Fax: 886-4-2287-7585; E-mail: sdyeh@nchu.edu.tw