Bot. Bull. Acad. Sin. (2002) 43: 271-276
Hou et al. SPTI degradation by aspartic type protease
An aspartic type protease degrades trypsin inhibitors, the
major root storage proteins of sweet potato (Ipomoea batatas (L.) Lam cv. Tainong 57)
Wen-Chi Hou1,*, Dong-Jiann Huang2, and Yaw-Huei Lin2,*
1Graduate Institute of Pharmacognosy Science, Taipei Medical University, Taipei 110, Taiwan
2Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan
(Received May 7, 2002; Accepted June 19, 2002)
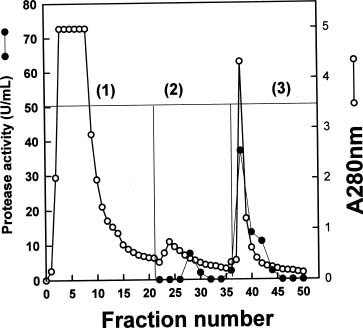
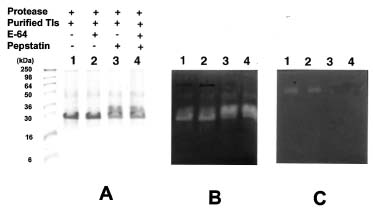
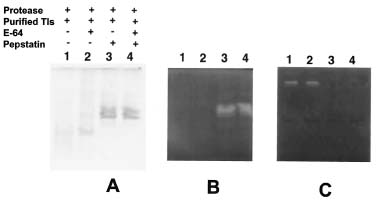
Abstract. Roots of sprouted sweet potato (Ipomoea batatas [L.] Lam) were used as materials to purify proteases which degraded trypsin inhibitors (Tis), the root storage proteins of sweet potato (SP). The commercial pepstatin-agarose (crosslinked, 6%) was chosen as an affinity column for purification. The purified protease has a molecular mass of about 64 kDa on the gelatin-SDS-PAGE gel and was inhibited by pepstatin but not by E-64 on the gelatin-SDS-PAGE gel. Therefore, it might belong to the aspartic type. Using the trypsin inhibitor activity staining method as a criterion for TI degradations, we found that this aspartic type protease could degrade purified Tis in the presence or absence of 5 mM DTT and the hydrolysis was complete in the former condition. The physiological role of aspartic type protease in the degradation of SPTis is discussed.
Keywords: Aspartic type protease; Degradation; Physiological role; Sweet potato; Trypsin inhibitor.
Abbreviations: SDS-PAGE, sodium dodecylsulfate-polyacrylamide gel electrophoresis; SP, sweet potato; TI, trypsin inhibitor.
Introduction
Proteases play important roles in post-translational modification, protein turnover, activation and inactivation of specific proteins, and nutrient supplementation (North, 1982). In plant tissues, specific proteases involved in the mobilization of reserve proteins (Chrispeels and Boulter, 1975; Wilson et al., 1986; Qi et al., 1992; Bottari et al., 1996; Senyuk et al., 1998; Davy et al., 2000), developmental processes (Lin and Tsai, 1991; Lin and Chan, 1992; Lin and Tsai, 1994; Dominguez and Cejudo, 1996; Voigt et al., 1997), and senescence (Hensel et al., 1993; Lohman et al., 1994; Smart et al., 1995; Drake et al., 1996) have been studied intensively.
Proteinaceous protease inhibitors in plants may be important in regulating and controlling endogenous proteases and in acting as protective agents against insect and/or microbial proteases (Ryan, 1973, 1989). Sohonie and Bhandarker (1954) reported for the first time the presence of trypsin inhibitors (TIs) in sweet potato (SP). Later, we indicated that TI activities in SP are positively correlated with concentrations of water-soluble protein (Lin and Chen, 1980), and that a large negative correlation exists between the natural logarithm of TI activities and cumulative rainfall, which suggests that SPTI activities may vary
in response to drought (Lin, 1989). Polyamines, including cadaverine, spermidine and spermine, were bound covalently to SPTI, which might participate in regulating the growth and developmental processes of SP (Hou and Lin, 1997a). SPTIs were also proved to have both dehydroascorbate reductase and monodehydroascorbate reductase activities and might respond to environmental stresses (Hou and Lin, 1997b). We found that TIs in SP roots accounted for about 60% of total water-soluble proteins and could be recognized as storage proteins (Lin and Chen, 1980). Maeshima et al. (1985) identified sporamin as the major storage protein in SP root, accounting for 80% of the total proteins there; however, a dramatic decrease to 2% of the original value was found during sprouting. Lin (1993) considered sporamin one form of TI in SP, a finding confirmed later by Yeh et al. (1997a). However, few reports concern the degradation of SP root storage protein during sprouting. In this work we report preliminary results showing that SP proteinaceous trypsin inhibitors were degraded by an endogenous aspartic type protease.
Materials and Methods
Plant Materials
Fresh roots of sweet potato (Ipomoea batatas L. Lam cv. Tainong 57) were purchased from a local wholesaler. After cleaning with water, the roots were either immediately cut into strips for Ti extraction according to the method of Hou and Lin (1997a) or placed in the
*Corresponding author. Prof. Yaw-Huei Lin: Fax: 886-2-2782-7954, E-mail: boyhlin@ccvax.sinica.edu.tw; or Prof. Wen-Chi Hou: Fax: 886-2-2378-0134, E-mail: wchou@tmu.edu.tw