Bot. Bull. Acad. Sin. (2002) 43: 277-282
Kao et al. Nutrient contents, d13C and d15N in mangrove
Nutrient contents, d13C and d15N during leaf senescence in the mangrove, Kandelia candel (L.) Druce
Wen-Yuan Kao1,*, Hung-Chieh Tsai1, Chen-Ning Shih1, Tyng-Tyng Tsai1, and Linda L. Handley2
1Institute of Botany, Academia Sinica, Taipei 115, Taiwan, Republic of China
2Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DQ, Scotland, UK
(Received July 2, 2001; Accepted May 9, 2002)
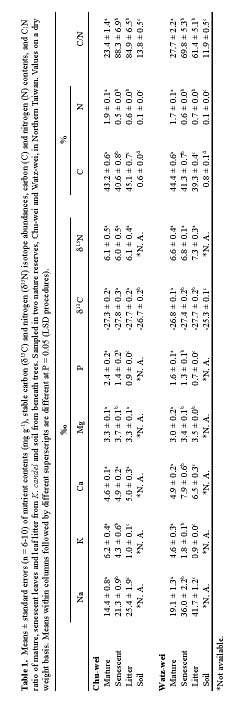
Abstract. Kandelia candel (L.) Druce is the dominant mangrove species in two nature reserves in Northern Taiwan. We characterized the nutrient quality of the leaf litter originating from K. candel and attempted to suggest the extent to which the observed decline of nutrient concentrations in senescing leaves could be attributed to the withdrawal of these nutrients into the main plant before abscission. On a dry weight basis, senescent leaves and leaf litter had significantly lower concentrations of N, P and K and higher concentrations of Na than did younger ones. At one site Ca and Mg contents rose with senescence; at the second site there was no change with age of leaf. Leaf d13C and C:N ratios suggest that withdrawal into the main plant may not be the major cause for the observed decline of C and N during leaf senescence. The d13C values of mature leaves ranged from -26 to -30; senescent leaves and leaf litter were 0.6 more negative for d13C than fresh leaves at the most seaward site. We attribute this decline of d13C to loss of stomatal control during senescence. d15N ranged from +4.3 to +9.8 for individual samples with no significant differences among ages of leaves, indicating no important gaseous N losses during senescence. P was probably not limiting to growth at these sites.
Keywords: 13-C; 15-N; Kandelia candel; Leaf litter quality; Mangrove.
Introduction
Mangrove ecosystems in tropical and subtropical coastal areas are highly productive, and mangrove litter is an important food sources for vertebrates and invertebrates (Robertson and Daniel, 1989; Osborne and Smith, 1990). Hence, an understanding of the nutrient contents of mangrove leaves can assist in understanding the quality of food resource available to consumers. It is additionally useful to know the fates of putative nutrient losses from mangrove. If, for instance, the nutrients are retained in the plant, they are unavailable to the larger ecosystem while the mangrove lives. If they are leached from leaves into the water, then they are available to other organisms, albeit not as a part of leaf litter.
Although N uptake in K. candel has been studied (Kao et al., 2001), the nutrient status of K. candel leaves during leaf senescence has not. We aimed to characterize the concentrations of leaf nutrients (C, N, P, K, Na, Ca and Mg) and to use d13C and d15N, in conjunction with elemental concentrations, to suggest whether decreases in leaf nutrient concentrations were related to withdrawal into the main plant.
Materials and Methods
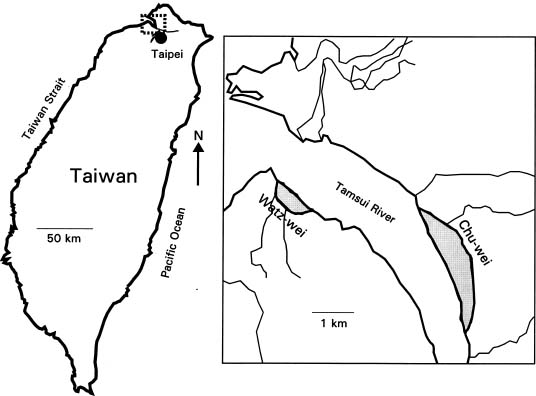
Leaves and soil samples were collected from two nature reserves (Figure 1), Watz-wei and Chu-wei swamps (25°9' N, 121°26' E), in May and June of 1998, respectively. Chu-wei swamp contains about 50 ha of pure K. candel forest and Watz-wei swamp about 9 ha. Tides are semidiurnal; salinity in Watz-wei Swamp is 5 to 30 psu and ranges from 1.1 to 27.4 psu in Chu-wei Swamp (Shae, 1995). Chu-wei lies further inland than Watz-wei and is closer to the sewage source from Taipei. The pH of water overlying soil at both sites is 7.1 to 7.9 (Shae, 1995). Mean annual temperature is 22°C, and mean annual precipitation is 2,100 mm (Climatological Data Annual Report, Central Weather Bureau, ROC.).
Mature leaves (the second or third pairs of leaves from the terminal bud) and senescent leaves (5 to 6 leaves per sample) were collected from each of 10 trees; leaf litter (as partially decomposed leaves; n = 5 to 6) was collected beneath each tree. Senescent leaves were selected as those which were yellow but still attached to the trees. A five-cm-deep core of the upper mineral soil was collected beneath each tree (n = 10) using a stainless steel hand corer. These were sealed in plastic bags on ice until analysis. Leaf samples were rinsed with distilled water; then leaf and soil samples were oven-dried (70°C) to a constant weight and ground to a fine powder with a mortar and pestle.
*Corresponding author. Phone: 886-2-2789-9590; Fax: 886-2-2782-7954; E-mail: bowykao@ccvax.sinica.edu.tw