Bot. Bull. Acad. Sin. (2002) 43: 283-290
Li et al. Pigment analysis and algal community investigations
Phytoplankton pigment analysis by HPLC and its application in algal community investigations
Hsiu-Ping Li1, Gwo-Ching Gong2, and Tung-Ming Hsiung1,*
1Institute of Bioscience and Biotechnology, National Taiwan Ocean University, 202 Keelung, Taiwan
2Department of Oceanography, National Taiwan Ocean University, Keelung, Taiwan
(Received May 29, 2001; Accepted May 28, 2002)
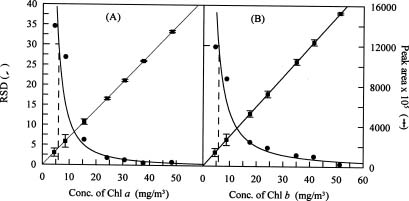
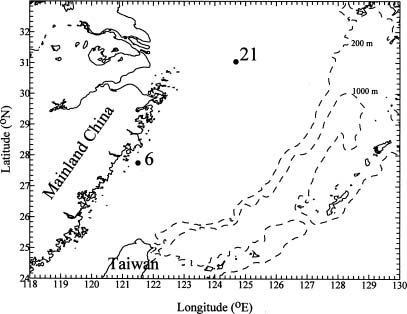
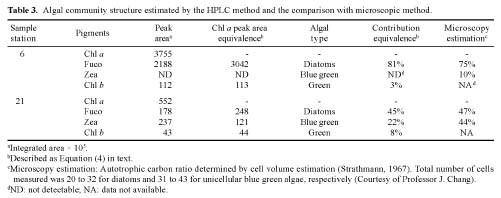
Abstract. A high performance liquid chromatography (HPLC) system for pigment analysis of marine phytoplankton was established. The characteristic quantification limits of the major photosynthetic chlorophylls a and b were 0.29 and 0.27 ng, respectively. The reliability of this method was verified by resolving the marker pigments in culture extracts from diatoms, blue-green algae and green algae. Fucoxanthin, zeaxanthin and chlorophyll b were selected for calculating the chlorophyll a/marker pigment ratios for the three types of algae, which were 1.39, 0.51 and 1.01, respectively. The composition of the phytoplankton community in summer samples, collected from the continental shelf of the East China Sea, was estimated by simple regression of these ratios. When results of composition abundance of diatoms and blue-green algae obtained by this method were compared with those obtained by the microscopic technique, the relative percentage difference was less than 6% for diatoms. On the other hand, a lower abundance of blue-green algae was estimated by the HPLC method.
Keywords: Algal community; HPLC; Marker pigments; Phytoplankton.
Introduction
Marine primary productivity can be referred to as the energy conversion generator for the marine food web. The abundance of marine phytoplankton in the oceans amounts to one-fourth of all plants in the world (Jeffrey and Hallegraeff, 1990). Phytoplankton, then, clearly play a critical role in global carbon fluxes (Falkowski et al., 1998). Chlorophyll a, the major photosynthetic pigment of marine phytoplankton, has been commonly used as an indicator of biomass or primary productivity in the ocean sphere. Various marine phytoplankton can be distinguished by their cellular diameter. For example, the cellular diameter of picoplankton is less than 2 µm, whereas that of nanoplankton or microplankton is larger than 20 µm. It has been reported that the carbon dioxide exchange flux in the oceans is significantly influenced by the size and assemblage of phytoplankton (Takahashi et al., 1993; Balch et al., 1996; Rivkin et al., 1996). Therefore, the distribution of, and variations in, the phytoplankton community, including in the chlorophyll contents, is critical in the study of marine carbon cycles.
Spectrophotometry and fluorometry are usually used to measure chlorophyll a (Lorenzen, 1967; Jeffrey and Humphrey, 1975). The former method is less sensitive and requires a large sample volume. Also, an appropriate empirical equation must be chosen (Jeffrey and Humphery, 1975; Porra et al., 1989). Despite having greater sensitivity, the fluorometric method tends to underestimate chloro
phyll a when chlorophyll b is unequally distributed in the water (Gieskes, 1991). Earlier investigations of these two methods have shown that their accuracy depended on the sample matrix and the calculation equations used (Lorenzen and Jeffrey, 1980; Wright and Jeffrey, 1997).
Thus far, most studies of phytoplankton communities have employed microscopy, using specific fixing agents for different algae. Pretreatment steps have included filtration and staining, and species identification and counting have been performed using light, fluorescence, or electron microscopy. Microscopy has been far from ideal, however. Obtaining data with high density and high frequency is somewhat difficult since the method requires experienced specialists and considerable time. Furthermore, some picoplankton, such as prochlorophyta, which possibly contributes to sea productivity, cannot be identified by microscopy (Chisholm et al., 1988). Flow-cytometric analysis has been developed for phytoplankton identification, but it must be based on a full understanding of the optical-characteristics of the species. Furthermore, flow-cytometry cannot be employed to measure large phytoplanktons (Jeffrey et al., 1999).
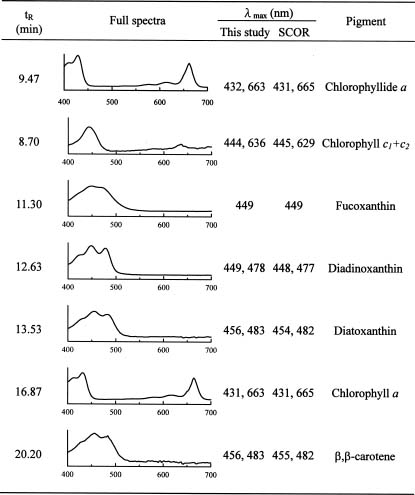
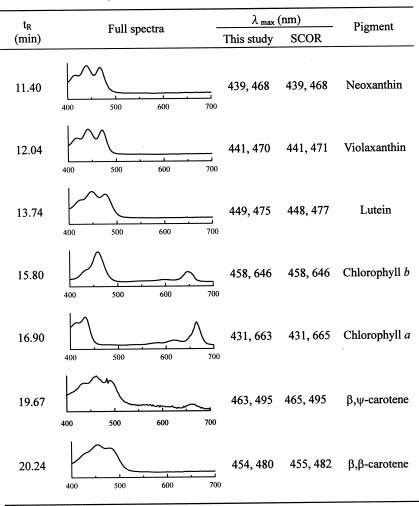
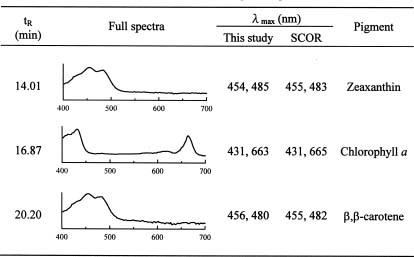
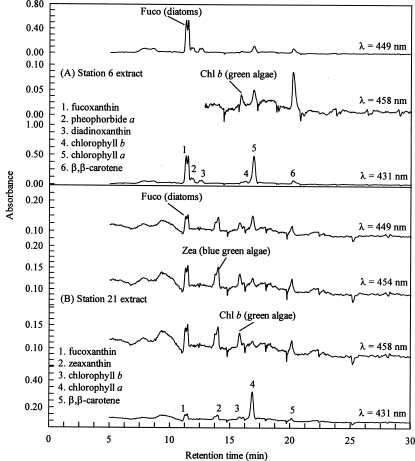
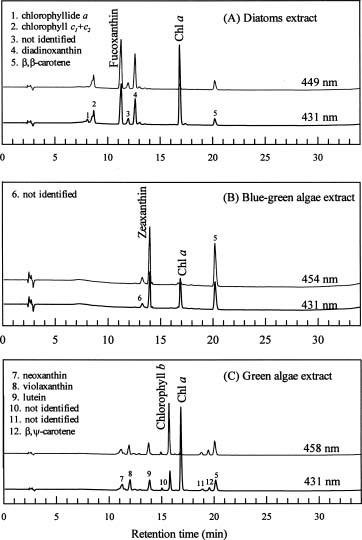
To overcome some of the inadequacies of microscopy, high performance liquid chromatography (HPLC) pigment method has been investigated in recent years to obtain both accurate chlorophyll a data and detailed information about the composition of phytoplankton communities (Mantoura and Llewellyn, 1983). This method is based on the premise that different algal classes have specific signature, or marker, pigments. For example, fucoxanthin, zeaxanthin, and chlorophyll b have been selected as taxonomical pigments for bacillariophyta (diatoms),
*Corresponding author. Tel: 886-2-24622192 ext. 5515; Fax: 886-2-24634243; E-mail: tmhsiung@mail.ntou.edu.tw