Bot. Bull. Acad. Sin. (2002) 43: 299-304
Yang et al. _ Allelopathic phenolics and chlorophyll accumulation
Effects of three allelopathic phenolics on chlorophyll accumulation of rice (Oryza sativa) seedlings: I. Inhibition of supply-orientation
Chi-Ming Yang, Chyoung-Ni Lee, and Chang-Hung Chou*
Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 11529, Republic of China
(Received November 1, 2001; Accepted May 2, 2002)
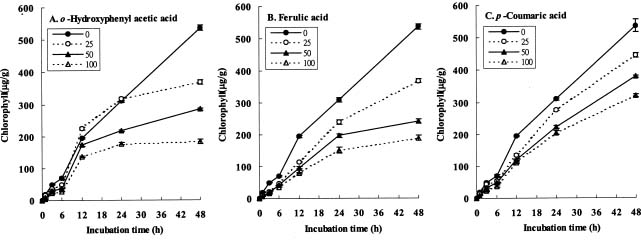
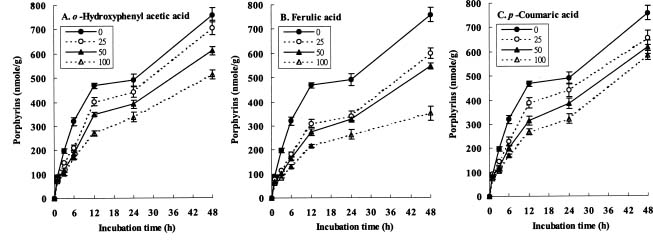
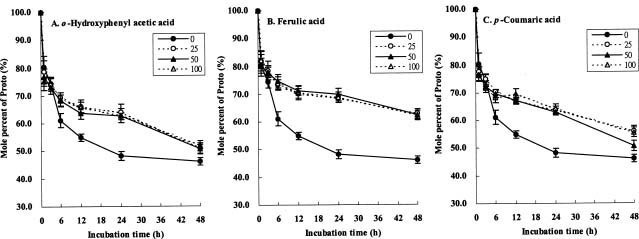
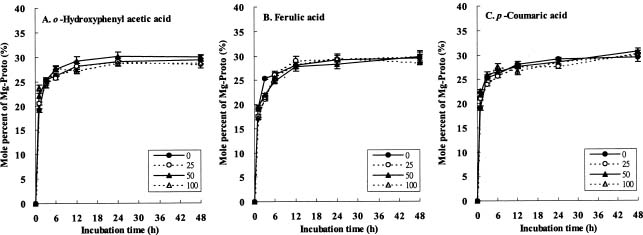
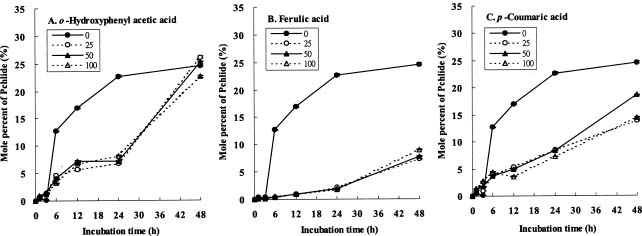
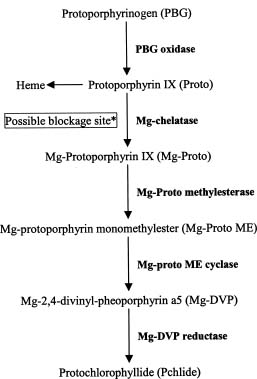
Abstract. The effects of three allelopathic chemicals, o-hydroxyphenyl acetic, ferulic, and p-coumaric acids, on the chlorophyll (Chl) supply-orientation in the rice leaf (Oryza sativa cul. TN67) were investigated. Two-week-old etiolated seedlings of rice were cultured in growth chamber in one-tenth Kimura's culture solution, with or without 25, 50 or 100 ppm of the phenolic compounds. The photoperiod, light intensity, and relative humidity were 12 h, 300 µmol/m2s, and 70%, respectively. Leaves were harvested at 1, 3, 6, 12, 24 and 48 h after treatment with phenolics. The concentrations of Chl and its three biosynthetic porphyrin precursors__i.e. protoporphyrin IX (Proto), Mg-protoporphyrin IX (Mg-Proto), and protochlorophyllide (Pchlide)__were determined, and the mole percent of total porphyrins was calculated. Accumulation of Chl and porphyrin contents were more inhibited as the phenolic concentrations increased. While the mole percent of Mg-Proto affected by the three allelochemical phenolics exhibited the same pattern, those of Proto and Pchlide showed quite different patterns of change in concentration. The data strongly suggest that Mg-chelatase may be the major target of the three phenolics. The order of inhibition strength is ferulic acid>p-coumaric acid>o-hydroxyphenyl acetic acid. It is concluded that the supply-orientation of Chl is significantly inhibited by the exogenous phenolics, causing the shortage of Chl.
Keywords: Allelopathic phenolic; Chlorophyll; Ferulic acid; o-hydroxyphenyl acetic acid; Oryza sativa; p-coumaric acid porphyrin; Supply-orientation.
Introduction
Allelopathy is the direct influence of chemicals released from one plant on the development and growth of another plant (Olofsdotter, 1998). It has been well documented for many years (Rice, 1984), but the mechanisms through which allelochemicals impact the development and growth of targeted plants remains obscure.
Multiple physiological effects have commonly been observed from treatments with many allelochemical phenolics. These effects include decreases in plant growth, absorption of water and mineral nutrients, ion uptake, leaf water potential, shoot turgor pressure, and osmotic potential caused by ferulic acid (Patterson, 1981; Einhellig et al., 1985; Gerald et al., 1992). Scopoletin, a coumarin derivative, inhibited dry matter production, leaf area expansion and photosynthesis in tobacco, sunflower and redroot pigweed (Einhellig et al., 1970). Chou and Lin (1976) found the aqueous extracts of decomposing rice residues in soil contained five phenolics and several unknown compounds and that extracts inhibited the radicle growth of lettuce and rice seeds and the growth of rice seedlings. Chlorogenic acid reduced stomatal aperture in tobacco and sunflower
(Einhellig and Kuan, 1971). Ferulic and p-coumaric acids reduced leaf water potential and stomatal diffusive conductance in sorghum and soybean (Einhellig et al., 1985). Einhellig (1995) proposed that a primary effect of phenolic acids is on the plasma membrane, and this perturbation contributes to a number of physiological effects causing growth reduction.
High levels of p-coumaric, ferulic, cinnamic and vanillic acids, and coumarin severely suppressed the photosynthesis of soybean and Lemna minor L. (Patterson, 1981; Einhellig, 1986). Three phenolic acids, p-coumaric and ferulic and vanillic acids, were also reported to severely inhibit photosynthesis and protein synthesis of isolated leaf cells of velvetleaf Abutilon theophrasti (Mersie and Singh, 1993). Chl reduction was observed in soybean plants treated with aqueous extract of velvetleaf (Colton and Einhellig, 1980) and in soybean and sorghum seedlings treated with p-coumaric and ferulic and vanillic acids (Einhellig and Rasmussen, 1979). It has also been suggested that some allelopathic compounds may interfere with the synthesis of porphyrin, precursors of Chl biosynthesis (Rice, 1984).
Even though a reduction of photosynthesis has been widely observed in the allelochemical-targeted plants, the component of photosynthesis which is directly or indirectly affected by the allelochemicals is still unknown. One possibility is that the allelochemicals may partially block
*Corresponding author. Tel: 886-2-2789-9590; Fax: 886-2-2782-7954; E-mail: chou@gate.sinica.edu.tw