Bot. Bull. Acad. Sin. (2003) 44: 13-18
Fu et al. RAPD analysis for genetic diversity in Changium smyrnioides
RAPD analysis for genetic diversity in Changium smyrnioides (Apiaceae), an endangered plant1
Chengxin Fu*, Yingxiong Qiu, and Hanghui Kong
Lab of Plant Systematic Evolution and Biodiversity, College of Life Sciences, Zhejiang University, Hangzhou 310029, P.R. China
(Received January 31, 2002; Accepted September 2, 2002)
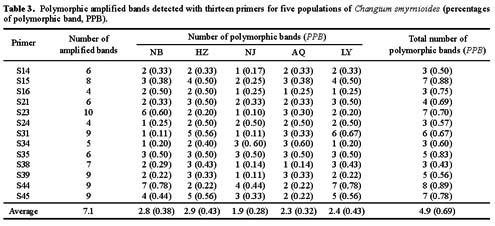
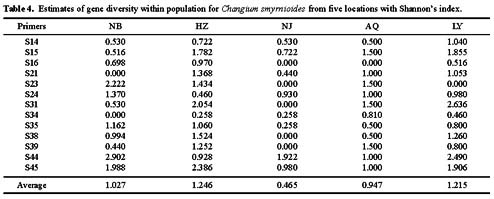
Abstract. Random Amplified Polymorphic DNA (RAPD) markers were used to measure genetic diversity of Changium smyrnioides Wolff (Apiaceae), a genus endemic to eastern China and an endangered medicinal plant, collected from five populations along the Yangzi River. A total of 92 amplified bands were scored from the 13 RAPD primers, and a mean of 7.1 amplified bands per primer and 69% (64 bands) percentages of polymorphic bands (PPB) was found. The Shannon's index was used to partition genetic diversity. Genetic diversity estimates indicated that 51.2% of total diversity was among populations and 48.8% within populations. The species shows higher genetic diversity between populations than other endangered plants. In this study, the RAPD results of genetic diversity are similar to those of previous studies employing allozymes (58% among populations and 42% within populations). The RAPDs provide a useful tool for assessing genetic diversity of rare, endemic species and for resolving relationships among populations. The results show that the genetic diversity of this species is high, possibly allowing it to more easily adapt to environmental variations. A pattern of greater variation among rather than within populations was observed with both markers. This distributive pattern of genetic variation of C. smyrnioides populations provides important baseline data for conservation and collection strategies for this species. It is suggested that most of the populations should be sampled in ex-situation protection so as to retain as much genetic diversity as possible.
Keywords: Allozymes; Changium smyrnioides; Genetic diversity; RAPD.
Introduction
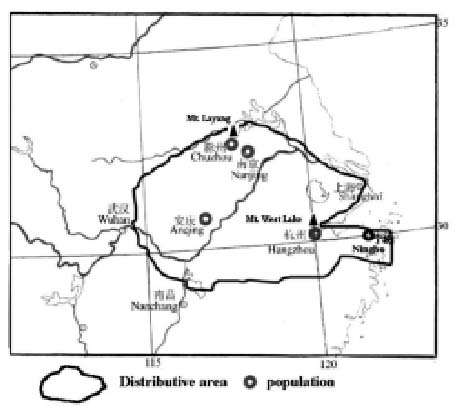
Changium smyrnioides Wolff, a species of monotypic genus in the family Apiaceae, is an endangered herbaceous perennial plant in China. It is endemic to eastern China, and is distributed only in fragments in Zhejiang, Anhui, Jiangsu, and Hubei Provinces (see Figure 1). The species is not only of important medicinal value; it is also of interest to plant systematists. The drugs made from this species promote the production of body fluid to quench thirst, nourish "yin" to moisten the lung, soothe the throat, and clear away toxic material that may cause skin infection. As there might be some internal reasons, such as questions of reproductive biology and population ecology, this species has been assigned an endangered status and is listed in the Red Data Book as an endangered plant by the government of China (Zhang, 1991).
The species was named C. smyrnioides by Wolff (1924) based on a specimen from Wuxin, Zhejiang. Various aspects of the genus have been discussed in earlier papers (Qiu et al., 2000; Qiu and Fu, 2001), but it should be emphasized that natural populations of C. smyrnioides are very small and rare. Because allozymes have sometimes
proven insufficiently variable for assessing genetic diversity within and among populations of rare endemic plants, attention has turned to more variable regions of the genome. A more recently employed approach in plant systematics and population biology is random amplified polymorphic DNA (RAPD) markers, a PCR-based (polymerase chain reaction) technique.
1State Key Basic Research and Development Plan of China, G2000046806, and project of NNSF of Zhejiang, China, No. 395064.
*Corresponding author. Tel: +86-571-86971576; Fax: 86-571-86971634; E-mail: fucxsmi@mail.hz.zj.cn
Figure 1. The distribution area and sampling locations of Changium smyrnioides.