Bot. Bull. Acad. Sin. (2003) 44: 43-52
Chang and Yang Microalgae for biofixation of carbon dioxide
Some characteristics of microalgae isolated in Taiwan for biofixation of carbon dioxide
Ed-Haun Chang1 and Shang-Shyng Yang*
Department of Agricultural Chemistry, National Taiwan University, Taipei, Taiwan 10617
(Received February 22, 2002; Accepted August 13, 2002)
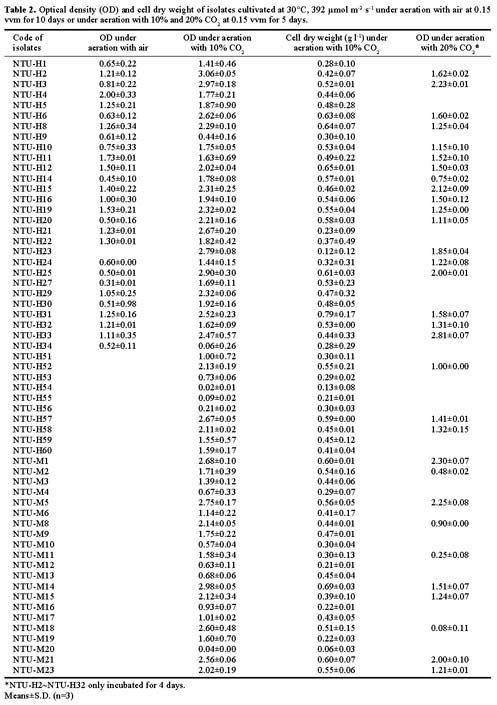
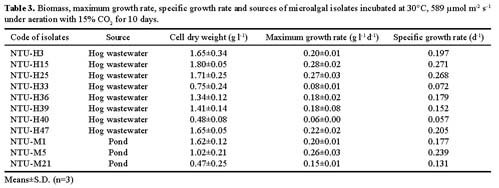
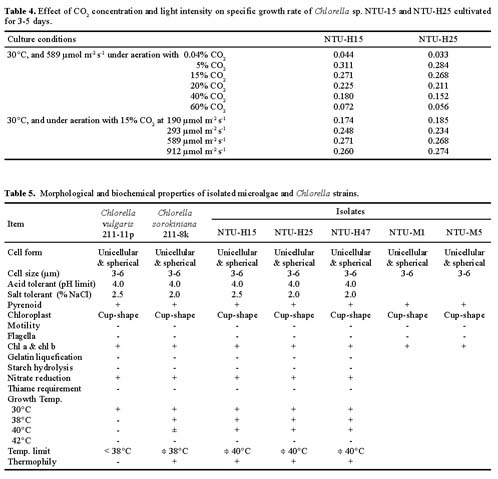
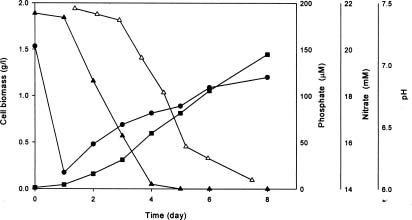
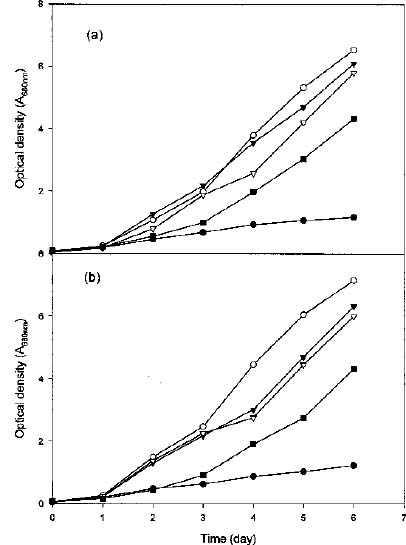
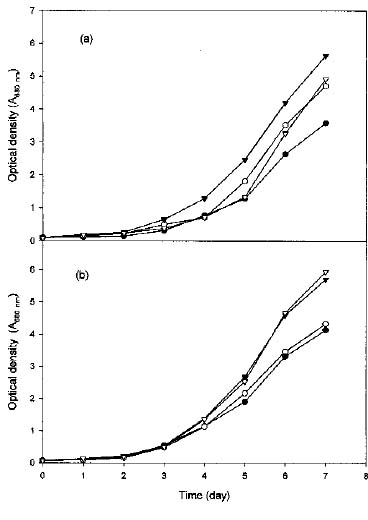
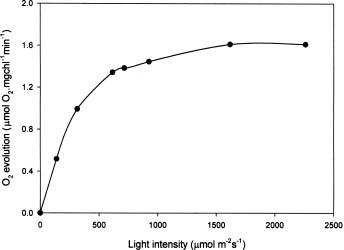
Abstract. To contribute the biological mean of CO2 fixation, more than 200 microalgal isolates were screened from lakes, ponds, sediments, hog wastewater, paddy fields, hot springs, and seawater in Taiwan. Two unicellular microalgae, Chlorella sp. NTU-H15 and Chlorella sp. NTU-H25, were isolated from hog wastewater. In the laboratory, they were able to grow up even in aeration containing CO2 up to 40% and have growth rates of 0.21 to 0.22 g dry wt l-1 d-1 at 20% CO2. Both algae had the same growth rate in the range from 5 to 40% CO2 and had a similar light response between 190 and 589 µmol m-2 s-1. Chlorella sp. NTU-H15 had a higher growth rate than Chlorella sp. NTU-H25 at pH 4.0 and ³35°C. Chlorella sp. NTU-H15 was able to tolerate high concentrations of CO2, high cell density, and a broad-range of temperature and pH. Each liter of Chlorella sp. NTU-H15 produced 1.8 g of dry cell. The maximum growth rate was 0.28 g dry wt l-1 d-1, and the specific growth rate was 0.27 d-1 at 15% CO2. Each mg of chlorophyll produced 1.6 mM O2 min-1 at 700 µmol m-2 s-1 at 30°C and 10 mM NaHCO3. While each liter of Chlorella sp. NTU-H25 produced 1.7 g of dry cell, the maximum growth rate was 0.27 g dry wt l-1 d-1, and the specific growth rate was 0.27 d-1. Both isolates are suitable for dense cultivation to fix CO2 directly and to produce cell biomass.
Keywords: Biofixation; Carbon dioxide; Chlorella; Growth rate and microalga.
Introduction
Global warming induced by increasing concentrations of greenhouse gases in the atmosphere is a matter of great environmental concern. Carbon dioxide is the principal greenhouse gas. Atmospheric CO2 has increased from 280 to 368 ppmv in the last 200 years and is responsible for about 50% enhancement in the greenhouse effect (Karube et al., 1992). Annual anthrophogenic emissions of CO2 are estimated to be 2 × 1010 tons, primarily from combustion of fossil fuels in association with an increasing population and industrialization. Recently, many attempts have been made to reduce atmospheric CO2. Physical and chemical treatments have been used to separate and recover CO2. Microalgal photosynthesis has increasingly received attention as a means of reducing the emission of CO2 into the atmosphere and producing industrially valuable compounds (Kodama et al., 1993; Kurano et al., 1995a; Lee et al., 1998; Yang et al., 2000).
Biofixation and utilization of CO2 by microalgae are among the most productive biological methods of treating industrial waste emissions, and the yield of biomass per acre is three to fivefold greater than from typical crops (Law and Berning, 1991; Akimoto et al., 1994). Direct use of flue gas reduces the cost of pretreatment, but the high concentration of CO2 and the presence of SOx and NOx inhibit the growth of cyanobacteria and microalgae
(Kurano et al., 1995b). A few works have recently reported the isolation of highly CO2-tolerant microalgae and cyanobacteria for biological fixation of CO2, such as Anacystis, Botryococcus, Chlamydomonas, Chlorella, Emiliania, Monoraphidium, Rhodobacter, Scenedesmus, Spirulina and Synechococcus (Hanagata et al., 1992; Takeuchi et al., 1992; Kodama et al., 1993; Sawayama et al., 1995; Takano and Matsunaga, 1995; Watanabe and Hall, 1995; Yamada et al., 1995; Zeiler et al., 1995; Ike et al., 1996; Yun and Park, 1997; Sung et al., 1999). The goal of this study is to isolate microalgae in Taiwan which can tolerate high CO2 concentrations and high temperatures in order to biofix carbon dioxide and discover the optimal conditions for biomass production.
Materials and Methods
Sources of Isolates
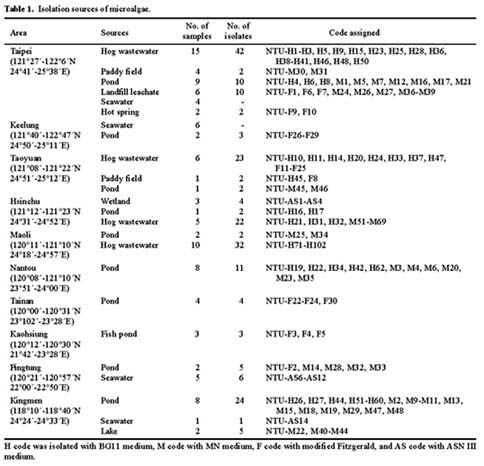
Microalgae were isolated from several samples taken from rivers, lakes, ponds, paddy fields, sediments, hog wastewater, ocean and hot springs in Taiwan (Table 1).
Culture Media
Medium BG-11 contained (g l-1): NaNO3, 1.5; K2HPO43H2O, 0.04; MgSO47H2O, 0.075; CaCl22H2O, 0.036; citric acid, 0.006, ferric ammonium citrate, 0.006; Na2EDTA, 0.001; Na2CO3, 0.02 and trace metal solution 1 ml (including H3BO3 2.86 g, MnCl24H2O 1.81 g, ZnSO47H2O 0.222 g, Na2MoO42H2O 0.390 g, CuSO45H2O 79 mg and Co(NO3)26H2O 49.4 mg per liter) at pH 7.4. Me
1Present address: Mackay Junior College of Nursing.
*Corresponding author. Tel: 886-2-23621519; Fax: 886-2-23679827; E-mail: ssyang@ccms.ntu.edu.tw