Bot. Bull. Acad. Sin. (2003) 44: 53-58
Barkosky and Einhellig Allelopathic interferenceof plant-water relationships
Allelopathic interference of plant-water relationships by para-hydroxybenzoic acid
Richard R. Barkosky1,* and Frank A. Einhellig2
1Minot State University, 500 University Avenue West, Minot, North Dakota 58707, USA
2Southwest Missouri State University, 901 South National Avenue, Springfield, Missouri 65804, USA
(Received April 9, 2002; Accepted August 15, 2002)
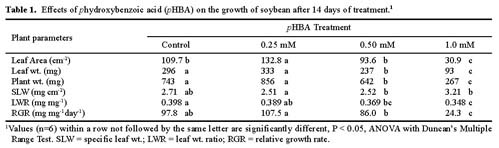
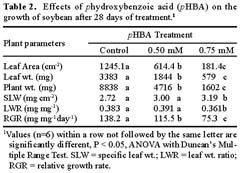
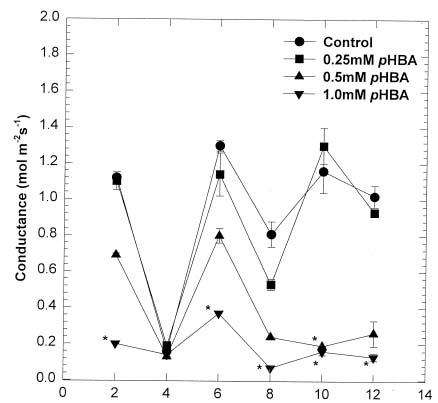
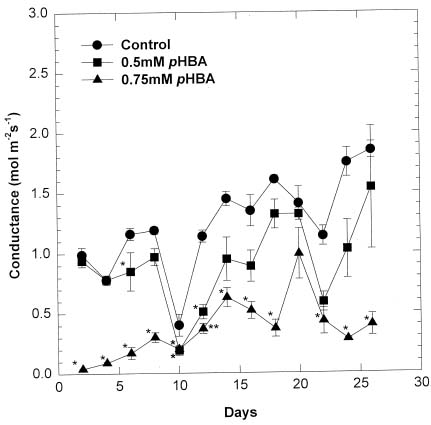
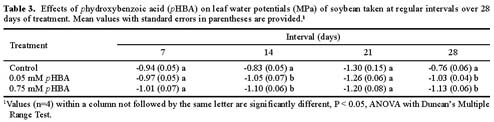
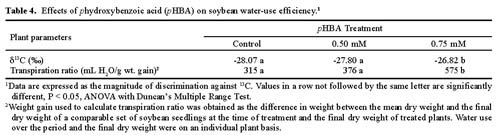
Abstract. Soybean seedlings were used as the test species to investigate the effect of phydroxybenzoic acid (pHBA) on growth and plant-water relationships. Plants were grown in nutrient solution under greenhouse conditions and were subjected to pHBA through amendments to the growth medium. Treatments were initiated 10 days after germination and continued for either 14 or 28 days with stomatal conductance, water potential, and water use monitored periodically. At harvest, effects on growth were determined and, in the 28-day study, the carbon isotope ratio (13C : 12C) of leaf tissue was analyzed as an indicator of integrated effects on plant-water status. Soybean growth was significantly reduced by 0.5 mM pHBA, or higher concentrations, with the degree of inhibition being concentration dependent. Plants treated with 0.75 mM pHBA had significantly lower stomatal conductance, lower water potential, and less discrimination against 13C. Similar trends were apparent in 0.5 mM pHBA-treated plants. Interference with plant-water balance appears to be one mechanism of action of pHBA causing a reduction in plant growth.
Keywords: Allelochemical; Allelopathy; Carbon isotopes; phydroxybenzoic acid; Soybean; Water status.
Introduction
The benzoic acid derivatives produced by higher plants have been frequently implicated in allelopathy (Rice, 1984). Para-hydroxybenzoic acid (pHBA; 4-hydroxybenzoic acid) has been isolated from a variety of crop residues and agricultural soils (Whitehead, 1964; Guenzi and McCalla, 1966; Chou and Patrick, 1976; Blum et al., 1991). It is one of several allelopathic Sorghum compounds found in aqueous extracts of plant material and associated with decomposing crop residues, germinating seeds, and root exudates (Abdul-Wahab and Rice, 1967; Hussain and Gadoon, 1981; Lehle and Putnam, 1983; Alsaadawi et al., 1986; Panasiuk et al., 1986; Einhellig and Rasmussen, 1989). Other plants implicated in allelopathic release of pHBA include Camelina alyssum and its influence on flax (Grummer and Beyer, 1960), several members of the genus Althaea (Gude and Bieganowski, 1990), and the grass Imperata cylindrica (Hussain and Abidi, 1991). Some of the reduction in root and coleoptile growth of wheat seedlings by wild oat (Avena fatua) root exudate is attributed to pHBA (Perez and Ormeno-Nunez, 1991).
Although several physiological effects of pHBA have been reported, its primary action affecting growth is obscure. Lee and Skoog (1965) found the hydroxybenzoic acids caused inactivation of indoleacetic acid. Effects on
respiratory metabolism have been suggested, as pHBA inhibited succinate hydrogenase and altered oxidation of NADH (Hulme and Jones, 1963; Lee, 1966). Glass (1973, 1974, 1975) demonstrated the inhibitory action of hydroxybenzoic acids on potassium and inorganic phosphate absorption of barley roots and related work established that membrane depolarization occurred (Glass and Dunlop, 1974). Stomatal effects from pHBA interaction with abscisic acid have been suggested (Purohit et al., 1991). Our previous work established that related phenolic compounds caused stomatal closure and other constraints on plant water relationships (Barkosky and Einhellig, 1993; Barkosky et al., 1999; Barkosky et al., 2000). Mechanism of action models proposed by Einhellig (1986) have suggested that membrane perturbations are probably a primary action of many phenolic allelochemicals.
This study was designed to determine the impact of pHBA on plant-water relationships and to evaluate the correspondence between these actions with effects on plant growth. As a part of the data collection, the relative concentration of stable carbon isotopes in leaf tissue was used as an indicator of water stress over time (Farquhar et al., 1982; Roeske and O'Leary, 1984; O'Leary, 1988; Berry, 1989; Tieszen, 1991). This analytical theory is based on the fact that several plant factors and environmental conditions, including water stress, contribute to the extent to which plants discriminate against carbon-13 (Farquhar et al., 1982, 1989; Ehleringer, 1989; Guy et al., 1989).
*Corresponding author. Telephone: 701-858-3116; Fax: 701-858-3163; E-mail: barkosky@minotstateu.edu