Bot. Bull. Acad. Sin. (2003) 44: 59-66
Wu et al. Naturalized Fabaceae (Leguminosae) species in Taiwan
Naturalized Fabaceae (Leguminosae) species in Taiwan: the first approximation
Shan-Huah Wu1, Shu-Miaw Chaw2, and Marcel Rejmánek1,*
1 Section of Evolution and Ecology, University of California at Davis, California 95616, USA
2 Institute of Botany, Academia Sinica, Nankang, Taipei, TAIWAN 11529
(Received July 23, 2002; Accepted October 24, 2002)
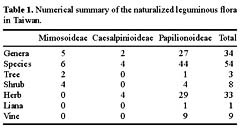
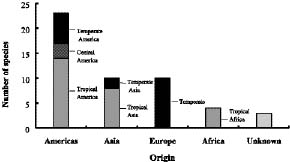
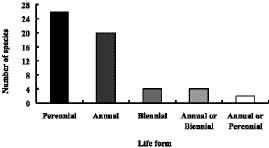
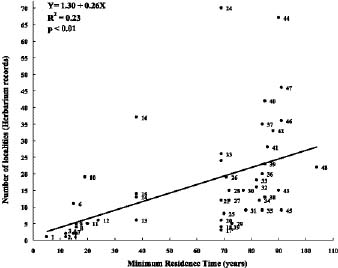
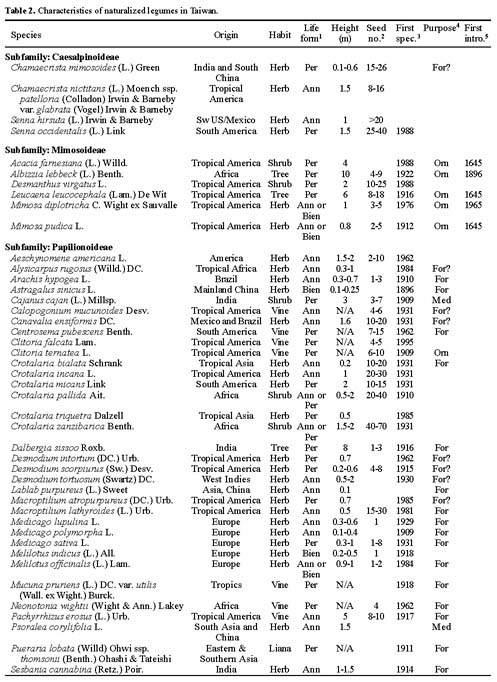
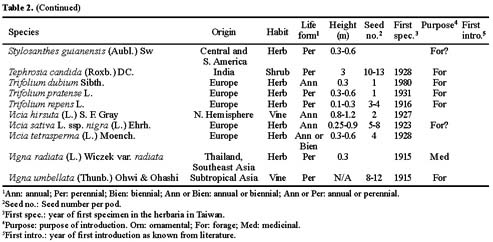
Abstract. The family Fabaceae (Leguminosae) is used as a starting point for creating a database of naturalized/invasive plant taxa in Taiwan, a Pacific island with very limited available information on plant invasions. A comprehensive list of naturalized Fabaceae, with background information on origin, habit, life form, and minimum residence time for most of the species is presented. This family is the second largest dicotyledon family in Taiwan, with non-native species accounting for one-sixth of the naturalized flora of this island: 54 naturalized species (approximately 23.8% of resident legume flora, including native and naturalized species) represent 34 genera (eleven exotic to Taiwan) and three subfamilies. Most of the naturalized taxa belong to the subfamily Papilionoideae, and the majority of them are herbaceous perennials with a minimum resident time ³ 60 years. Among these naturalized species, two are listed among the world's worst weeds, and at least six of them are known as important environmental weeds in other parts of the world. Most of the naturalized legumes were introduced from the Neotropics, and a notable portion (at least 50%) of them was introduced deliberately for cultivation as forage or ornamentals. Naturalized legumes often exhibit different rates of spread within similar time frames. For example, Crotalaria zanzibarica is known from approximately 70 localities, while C. bialata is known from only three different localities even though both have the same minimum residence time of 69 years. Such differences suggest that different taxa have a different degree of invasiveness.
Keywords: Alien flora; Habit; Invasive; Life form; Minimum residence time; Pacific island.
Introduction
Plant invasions have been recognized as one of the most serious global processes impacting the structure, composition, and function of natural and semi-natural ecosystems (Mooney and Hobbs, 2000; Vitousek et al., 1997). Ecologists are now paying more attention to plant invasions, including their history and impacts. Several conclusions, concepts, and hypotheses have been put forward and evaluated, especially those related to biogeographical settings, historical events, and biological traits. For example, it has been shown that a species that invades one place tends to be invasive also in other areas (Meyer, 2000); some characteristics possessed by particular groups of species can be used to predict their invasiveness (Reichard and Hamilton, 1997; Rejmánek and Richardson, 1996); and plants of different geographical origin seem to differ in their success (Huston, 1994).
Despite the recent recognition of the importance of invasive species, there are still many areas of the world where basic information on naturalized plant taxa and plant invasions is only anecdotal or completely lacking. Taiwan, a tropical and subtropical Pacific island, is a good example:
only occasional and non-ecological attention has been paid to naturalized species on this island (e.g., Peng et al., 1998; Peng and Yang, 1998; Chen et al., 1999; Kuoh and Chen, 2000; Chen and Wu, 2001; Yang, 2001).
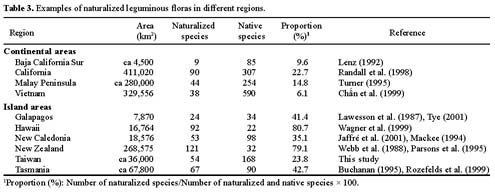
Among resident plant families, Fabaceae (Leguminosae) was chosen as a model for evaluation of plant invasions in Taiwan. The legume family is one of the largest vascular plant families and is the most notorious contributor to the naturalized flora of the world (Binggeli, 1996; Pyek, 1998). Approximately one-fourth of resident legumes in Taiwan are alien species. Furthermore, this family has been studied extensively by many taxonomists, and therefore, relatively comprehensive data are available and abundant specimen collections are accessible in local herbaria.
The goals of this study were: (1) to compile a comprehensive list of naturalized Fabaceae taxa in Taiwan; and (2) to summarize their basic biological characteristics and geographical sources.
Terminology
A "naturalized" species is defined as an introduced (non-native, exotic) species that can consistently reproduce and sustain populations over many generations without (or in spite of) direct intervention by humans. After successful local establishment, some naturalized species disperse and produce viable offspring in regions distant
*Corresponding author. Tel: 1-530-752-1092; Fax: 1530-752-1449; E-mail: mrejmanek@ucdavis.edu