Bot. Bull. Acad. Sin. (2003) 44: 99-105
Yan et al. Variation in camptothecin content
Variation in camptothecin content in Camptotheca acuminata leaves
Xiu-Feng Yan1,*, Yang Wang1, Tao Yu1, Yu-Hong Zhang2, and Shao-Jun Dai1,3
1Faculty of Forest Resources and Environment, Northeast Forestry University, Harbin 150040, P.R. China
2Key Laboratory for Forest Plant Ecology of the Ministry of Education, Northeast Forestry University, Harbin 150040, P.R. China
(Received April 9, 2002; Accepted November 5, 2002)
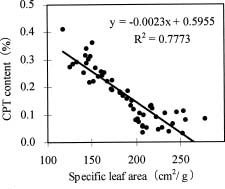
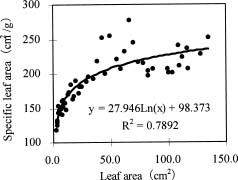
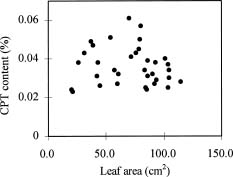
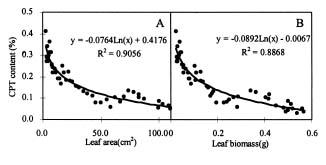
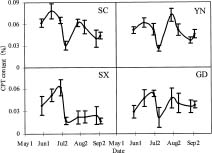
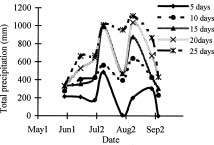
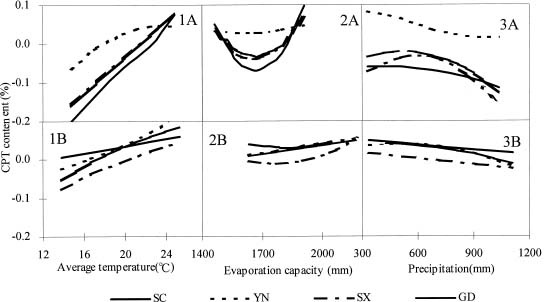
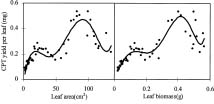
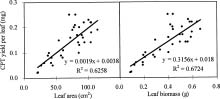
Abstract. Variations in camptothecin (CPT) content in relation to leaf characteristics namely leaf area, leaf biomass, and specific leaf area (leaf area divided by leaf biomass), and the influence of climatic factors on CPT content in saplings of Camptotheca acuminata Decne. were studied. Leaf area and CPT content showed a logarithmic relationship in young leaves similar to that between leaf biomass and CPT content, while CPT content was linearly correlated to specific leaf area. There was no apparent relationship between each leaf characteristic and CPT content in old leaves. CPT contents in sapling leaves showed large variation from May to September and were highly correlated to the variation in the level of climatic factors, where high average temperature, high evaporation capacity and low precipitation increased CPT content according to stepwise regression analysis. This suggests that adverse growing conditions can induce CPT accumulation to enhance chemical defense in C. acuminata. Young leaves with area of 70 cm2-100 cm2 gave the highest quantity of CPT per leaf, thus the optimal harvest period would be when young leaves expanded to these sizes. In addition, sample collection on dry days is preferable as it increases CPT content in raw materials.
Keywords: Camptothecin; Camptotheca acuminata leaves; CPT optimization.
Introduction
Camptothecin (CPT) is a monoterpenoid indole alkaloid originally isolated from Camptotheca acuminata Decne, a deciduous tree native to south China, that has gained great attention for its significant antitumor activities in experimental studies (Wall et al., 1966). Irinotecan (CPT-11) (Masuda et al., 1992; Abigeres et al., 1995; Bleiberg, 1999) and topotecan (TPT) (Lilenbaum et al., 1995; Romanelli et al., 1998; Clements et al., 1999), two water-soluble derivatives of CPT, have gained approval by the Food and Drug Administration of the United States of America (FDA) for treating colorectal and ovarian cancer. Other camptothecinssuch as 9-aminocamptothecin (9AC), 9-nitrocamptothecin (9NC), and 7-(4-methyl piperazino-methylene)-10,11-ethylenedioxycamptothecin (GG211)have also showed remarkable potential in the treatment of carcinoma (Wall and Wani, 1996; Giovanella, 1997; Jeha et al., 1998; Stevenson et al., 1999). Camptothecins are lauded as one of the most promising anticancer drugs of the twenty-first century (Li and Adair, 1994).
CPT acts as a defensive chemical in C. acuminata though there is not much research on this aspect of CPT. It was reported that goats feeding on C. acuminata leaves
became poisoned (Cao et al., 1986) as well as honeybees (Xia, 2000). In addition, CPT is strongly toxic to other animals. CPT is water-insoluble, but it and related analogs contain an a-hydroxy-d-lactone ring functionality which hydrolyzes under certain conditions, i.e. at pH 7 or above, with the lactone moiety readily opening up to yield the water-soluble carboxylate form (Burke, 1996) which is highly toxic to animals and humans. Extensive toxicological and pharmacological studies in mice, dogs, and pigs determined that CPT is toxic due to inflammatory ileitis and myelo suppression (Giovanella, 1997). Therefore, CPT plays a role in deterring herbivore feeding in C. acuminata. Thus understanding the variation in CPT content in C. acuminata will increase our knowledge of the regulation of chemical defenses, as well as help to optimize harvest opportunities for the raw materials of this important anti-cancer pre-drug.
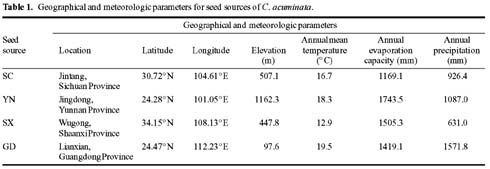
CPT is present in all organs of C. acuminata (Lin et al., 1977). Traditionally, CPT has been extracted from root, root bark, and fruits (Lin et al., 1977), but recent results have indicated that CPT exists at high levels in very young leaves, even higher than in the fruits (Lopez-Meyer et al., 1994; Zhang and Yang, 1997). In this paper, the production of CPT is related to leaf characteristicsnamely leaf area, leaf biomass, and specific leaf area (leaf area divided by leaf biomass)and the influence of climatic factors in CPT content are all investigated. Additionally, based on the findings, an optimal scheme for harvesting of C. acuminata leaves for CPT extraction is discussed.
3Ph.D. Candidate, working at Department of Biology, Harbin Normal University, Harbin, 150080, P.R. China.
*Corresponding author. Tel: +86-451-2192185; Fax: +86-451-2165504; E-mail: yanxf@public.hr.hl.cn