Bot. Bull. Acad. Sin. (2003) 44: 107-112
Wang & Yeh d13C of marine macroalgae from Taiwan
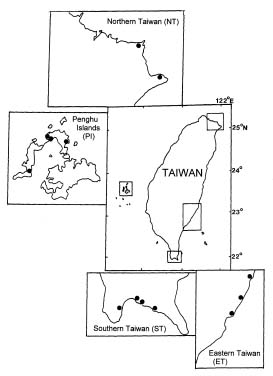
d13C values of marine macroalgae from Taiwan
Wei-Lung Wang1 and Hsueh-Wen Yeh2,*
1Department of Biology, National Changhua University of Education, Changhua 500, Taiwan
2Institute of Earth Sciences, Academia Sinica, Taipei 115, Taiwan
(Received April 18, 2002; Accepted January 10, 2003)
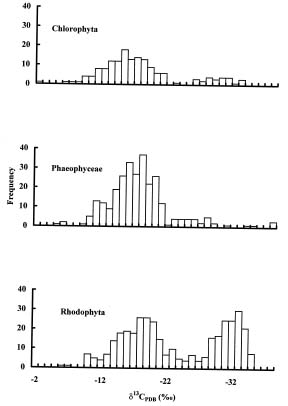
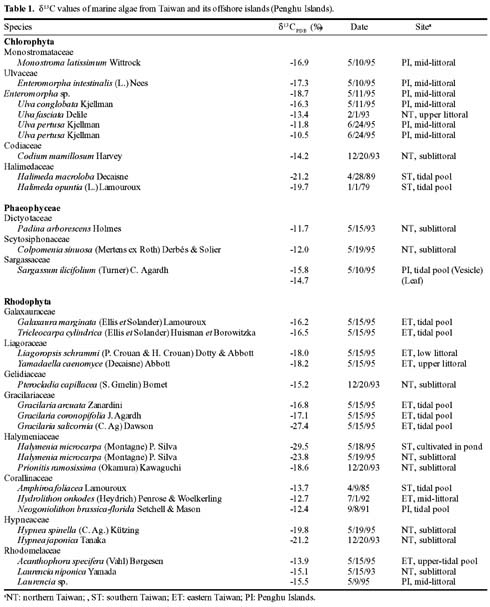
Abstract. The natural abundance of stable carbon isotope values (d13C) of organic matter of marine macroalgae collected from Taiwan and its offshore islands (Penghu Islands) were analyzed in this study. These values ranged from -10.5 to -29.5. The highest d13C value came from green algae (-10.5), Ulva pertusa while most algae exhibited values in the -14 to -19 range. On average, the d13C values of red algae (-17.7) are slightly more negative than those of green (-16.5) and brown (-13.6) algae. The carbon isotope record from organic matter of marine algae and its implications for marine algae physiology are also discussed.
Keywords: Marine macroalgae; d13C; Taiwan.
Introduction
Carbon isotope fractionation is associated with photosynthesis, and non-photosynthetic physical-chemical processes can also discriminate, e.g. lipid synthesis from carbohydrates (Farquhar et al., 1989). The carbon isotope composition of plants is employed to indicate photosynthetic pathways in terrestrial plants (Bender, 1968; Troughton, 1979). The d13C values for terrestrial C3, C4, and CAM plants range from -22 to -38, -11 to -19, and -13 to -34, respectively. The lower d13C values in the range for CAM plants refers to facultative CAM plants functioning as C3 plants. Carbon isotope combinations measured in marine algae range between -8.8 and -34.7, potentially leading to the mistaken impression that both C3 and C4 photosynthetic pathways are present (Raven et al., 1982; Stephenson et al., 1984; Dunton and Schell, 1987).
Many studies on the pathway of CO2 assimilation during photosynthesis in plants have demonstrated that differences arise in metabolism of the carbon isotopes 13C and 12C (Park and Epstein, 1960, 1961; Bender, 1968, 1971; Tregunna et al., 1970; Smith and Epstein, 1971; Troughton, 1979; Farquhar et al., 1989; Sackett, 1991; Johnston and Raven, 1992; Maberly et al., 1992; Raven, 1993). With terrestrial C3 and C4 plants, the degree of discrimination against 13CO2 is distinctive and falls in two clearly separable ranges (Bender et al., 1973; Smith et al., 1973; Yeh and Wang, 2001). The ranges for both C3 as well as C4 plants can largely be explained by variations in d13C values of source CO2 and/or in environmental conditions, in addition to probable species effect (Yeh and Wang, 2001).
However, with plants which assimilate CO2 via the CAM pathway, the extent of discrimination stretches over the entire range covering both in C3 and C4 plants (Bender et al., 1973; Osmond et al., 1973). Other work has shown that much of the isotope discrimination during photosynthetic CO2 uptake is by ribulose-bisphosphate carboxylase-oxygenase (RUBISCO) in terrestrial C3 plants (Park and Epstein, 1961; Whelan et al., 1973) while the carboxylation catalyzed by phosphoenolpyruvate carboxylase (PEPCase) shows only a slight isotopic discrimination in terrestrial C4 plants (Whelan et al., 1973). Since these are the two known carboxylases involved in photosynthetic CO2 assimilation, CO2 assimilation can be assessed via PEPCase or RUBISCO or both in a given tissue by studying the d13C value of the tissue. Reiskind and Bowes (1991) report that phosphoenolpyruvate carboxykinase (PEPCK) fixes CO2 in the "C4-like" marine green algae Udotea flabellum, and are involved in the high rates of dark CO2 fixation in diatoms and brown algae. Thus, an index of CO2 assimilation in an organism can be theoretically obtained by knowing the d13C value.
For subtidal algae two carbon sources are available for photosynthesis: CO2 and HCO3-. The marine macroalgae with d13C values lower than -30 are mainly subtidal red algae, with some shaded intertidal red algae and a few green algae, and those examined in the laboratory rely on diffusive CO2 entry (Raven, 1997; Raven et al., 2002). Some red algae with very low d13C values and diffusive CO2 entry live in warmer, high intertidal habitats, with significant CO2 gain from the atmosphere. Marine benthic organisms with very positive d13C values are mainly green macroalgae and seagrass, with some red and brown macroalgae, with a high d13C that can be accommodated by CO2 use without discrimination in favor of d13C (Raven et al., 2002). All of these organisms are able to use HCO3-.
*Corresponding author. Tel: 886-2-2783-9910 ext. 619; Fax: 886-2-2783-9871; E-mail: hwyeh@earth.sinica.edu.tw