Bot. Bull. Acad. Sin. (2003) 44: 123-128
Wang et al. An NLS in LLA23 for nuclear targeting
The nuclear localization signal of a pollen-specific, desiccation-associated protein of lily is necessary and sufficient for nuclear targeting
Huei-Jing Wang1, Guang Yuh Jauh2, Yau-Heiu Hsu1, and Co-Shine Wang1,*
1Graduate Institute of Biotechnology, National Chung Hsing University, Taichung, Taiwan 40227
2Institute of Botany, Academia Sinica, Taipei, Taiwan 11529
(Received August 22, 2002; Accepted November 26, 2002)
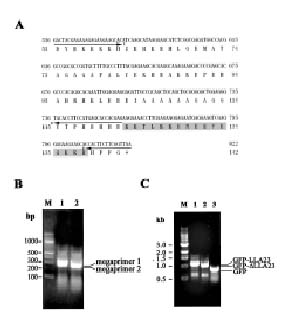
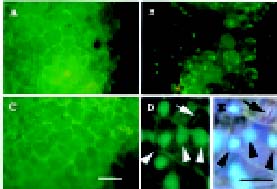
Abstract. The lily LLA23 protein represents a novel member of water-deficit/ripening-induced protein family (Plant Cell Physiol. 41: 477-485, 2000). Examination of the C-terminal half of LLA23 reveals the presence of basic regions that are reminiscent of a nuclear localization signal (NLS). To investigate the nuclear targeting property of NLS in LLA23, a green fluorescent protein (GFP) gene fused with the C-terminal half of LLA23 (GFP-LLA23) was constructed. In addition, the GFP alone and a mutGFP-DLLA23 that had the sequence for the putative NLS deleted were also constructed. All these three constructs were separately inserted into a bamboo mosaic potexvirus (BaMV) vector. Infection of BaMV in Chenopodium quinoa caused local lesions in leaves where the green fluorescence of fusion proteins could be visualized by fluorescence microscopy. The RNA blot and immunoblot analyses of BaMV-infected leaves indicated that the recombinant subgenomic RNA and the resulting protein were strongly detected. Fluorescence microscopic studies revealed that the NLS in LLA23 exhibited a property of nuclear targeting, showing highly condensed spots of green fluorescence in leaf cells, whereas GFP alone was apparently distributed throughout the cytoplasm. In contrast, a deletion of the NLS sequence resulted in exclusively cytoplasmic localization of the fusion protein. The nuclear location of the GFP-LLA23 protein in leaf cells was further confirmed by staining with 4´,6-diamidino-2-phenylindole. These results clearly demonstrate that the putative NLS in LLA23 is necessary and sufficient for import of the LLA23 protein into the nucleus.
Keywords: BaMV; Desiccation; Lilium longiflorum; NLS; Pollen-specific protein.
Abbreviations: BaMV, bamboo mosaic potexvirus; DAPI, 4´,6-diamidino-2-phenylindole; GFP, green fluorescent protein; Lea, late embryogenesis abundant; NLS, nuclear localization signal; PAGE, polyacrylamide gel electrophoresis; PBS, phosphate buffer saline; PCR, polymerase chain reaction.
Introduction
Pollen plays a crucial role in sexual reproduction of flowering plants. Within the anther, pollen mother cells undergo meiosis to produce a tetrad of haploid microspores that subsequently devide mitotically to form mature pollen grains. When pollen grows to full maturity, it exhibits various degrees of desiccation prior to anthesis and this desiccation process represents the last stage of pollen maturation.
The quantitative and qualitative changes in protein accumulation during pollen development have been documented in various plant species (Zarsky et al., 1985; Vergne and Dumas, 1988; Detchepare et al., 1989; Bedinger and Edgerton, 1990; Zarsky et al., 1995), but there have been few reports on pollen-specific proteins that accumulate in the maturing pollen. We have previously described the ac
cumulation of a set of newly synthesized anther/pollen-specific proteins prior to anthesis during anther development in lily (Wang et al., 1992; Wang et al., 1996). Ueda and Tanaka (1995) have found two male gamete-specific histone variants in the generative nuclei of lily pollen. Recently, pollen-specific proteins were also described in pollen of tobacco and lily (Wittink et al., 2000; Ko et al., 2002; Mogami et al., 2002).
We have characterized a number of pollen-specific proteins related to dehydration in lily pollen (Wang et al., 1996, 1999; Ko et al., 2002). Of these, the LLA23 protein represents a novel member of the water-deficit/ripening-induced protein family (Wang et al., 1998; Huang et al., 2000). The water-deficit/ripening-induced proteins reported in various plant species possess a putative nuclear localization signal (NLS) at the C-terminus (Iusem et al., 1993; Canel et al., 1995; Silhavy et al., 1995; Chang et al., 1996; Schneider et al., 1997). Subcellular fractionation experiments performed by Iusem et al. (1993) indicated that the Asr protein is located primarily in the nucleus.
*Corresponding author. Tel: 886-4-287-4754 ext. 105; Fax: 886-4-286-1905; E-mail: cswang2@nchu.edu.tw