Bot. Bull. Acad. Sin. (2003) 44: 129-140
Li et al. Over-expression of AtHsfA1b enhanced chilling tolerance
Over-expression of Arabidopsis thaliana heat shock factor gene (AtHsfA1b) enhances chilling tolerance in transgenic tomato
Hsiao-Yuan Li1, Chiung-Swey Chang1, Liang-Suei Lu1, Chien-An Liu2, Ming-Tsair Chan1,*, and Yee-Yung Charng1,*
1Institutes of BioAgricultural Sciences, Academia Sinica, Nankang, Taipei, Taiwan 115, R.O.C.
2Asian Vegetable Research and Development Center, Tainan, Taiwan 741, R.O.C.
(Received September 2, 2002; Accepted November 8, 2002)
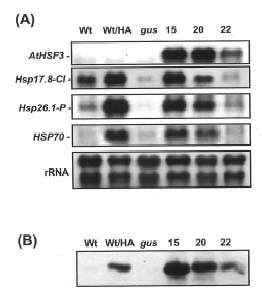
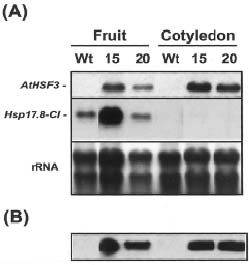
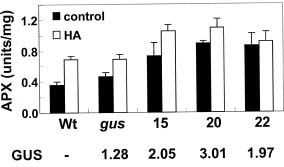
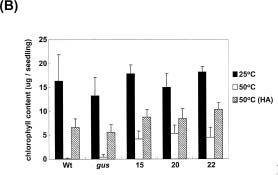
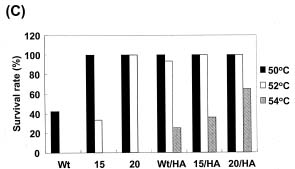
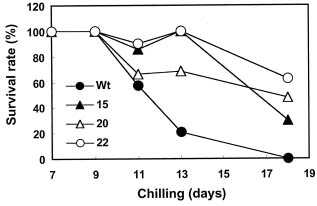
Abstract. Heat shock pre-treatment or heat acclimation can enhance the subsequent chilling tolerance of many plant species. However, the exact mechanism is currently unknown. Transgenic tomato plants harboring an Arabidopsis thaliana HsfA1b (AtHsfA1b) and b-glucuronidase (gusA) fusion gene under the control of a constitutive CaMV35S promoter were generated to evaluate whether heat shock factor (Hsf), a major transcription regulator of heat shock response, is involved in heat shock-induced chilling tolerance. The transcripts or protein product of heat-shock induced genes in the transgenic plants highly expressing AtHsfA1b-gusA accumulated to higher levels than those of the wild-type or gusA transgenic plants under non-heat shock conditions. This suggests that AtHsfA1b-GUS can cross-talk with the heat shock responsive machinery in transgenic tomato plants. In addition, the specific activity of soluble isoforms of ascorbate peroxidase of the AtHsfA1b-gusA transgenic plants was about twofold higher than that of the wild-type or gusA transgenic plants under non-heat stress conditions. Without heat acclimation, seedlings of the AtHsfA1b-gusA transgenic tomato lines showed a significantly higher level of thermal and chilling tolerance than that of the wild-type or gusA transgenic plants. Based on these results, we suggest that Hsf may play a pivotal role in heat-shock-induced chilling tolerance, and constitutive expression of the transcription regulatory gene in chilling sensitive crops may be useful in improving tolerance against chilling stress.
Keywords: Chilling stress; Heat shock factor; Transgenic tomato.
Abbreviation: HSICT, heat-shock-induced chilling tolerance; Hsf, heat shock factor; Hsp, heat shock protein; sHsp, small Hsp; GUS, b-glucuronidase;
Introduction
Chilling injury is a physiological disorder that develops in certain plants exposed to nonfreezing temperatures below about 12°C (Lyons, 1973; Saltveit and Morris, 1990). Many important crops indigenous to the tropics and subtropicssuch as banana, mango, papaya, rice, and tomatoare chilling sensitive (Saltveit and Morris, 1990). Chilling injury manifests a range of visible symptoms, often used as indicators of its severity, including reduced growth vigor, abnormal ripening, stimulated respiration and ethylene production, and increased cellular membrane leakage and disease susceptibility (Lyons, 1973; Saltveit and Morris, 1990). These symptoms can be reduced by heat shock treatment before exposure to chilling temperatures (Lafuente et al., 1991; Lurie and Klein, 1991; Saltveit, 1991; McCollum et al., 1993; Collins et al., 1995; Woolf et al., 1995; Rab and Saltveit, 1996; Sato et al., 2001) in a phenomenon termed heat-shock-induced chilling tolerance (HSICT). De
novo protein synthesis seems necessary for HSICT in mung bean hypocotyls (Collins et al., 1995). The exact mechanism of HSICT is so far unknown.
The induction of genes encoding heat shock proteins (Hsps) is one of the most prominent responses at the molecular level of organisms exposed to high temperature (Kimpel and Key, 1985; Lindquist, 1986; Vierling, 1991; Waters et al., 1996). According to their approximate molecular mass in kDa, the major eukaryotic Hsps have been grouped into five conserved classes: namely Hsp100, Hsp90, Hsp70, Hsp60, and the small Hsps (~16 to 42 kDa). Plant small Hsps (sHsps) can be further divided into at least six gene families encoding several proteins localized to the cytosol and nucleus (class I, II and III) and different organelles (chloroplast, endoplasmic reticulum, and mitochondria) (Scharf et al., 2001). Accumulation of Hsps increases thermotolerance and protects cells from the detrimental effect of high temperature, probably through their functioning as molecular chaperones (Waters et al., 1996). The correlation of HSICT and the accumulation of Hsps in plant tissues have been recently characterized (Collins et al., 1995; Sabehat et al., 1996; Kadyrzhanova et al., 1998). However, the role of Hsps, if any, in HSCIT is unclear.
*Corresponding authors. Fax: 886-2-2651-5600; E-mail: yycharng@gate.sinica.edu.tw, mbmtchan@ccvax.sinica.edu.tw