Bot. Bull. Acad. Sin. (2003) 44: 151-158
Lin et al. Regulation of circadian protein degradation
Factors affecting the circadian degradation of COP23 in Synechococcus RF-1
Rong-Fong Lin, Kun-Da Tsai, and Tan-Chi Huang*
Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 115, Republic of China
(Received October 3, 2002; Accepted February 11, 2003)
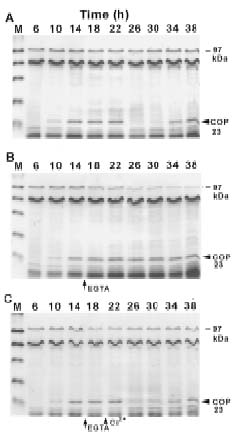
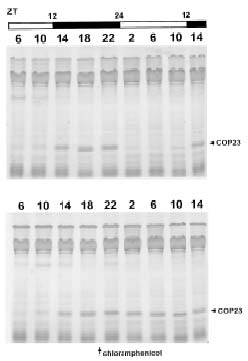
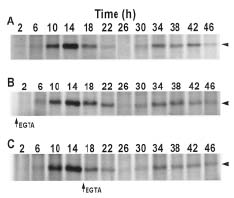
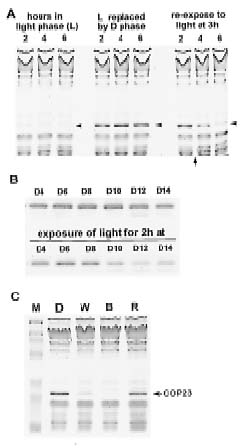
Abstract. The circadian rhythm of COP23, a protein located in the cell membrane of Synechococcus RF-1, was regulated by a circadian synthesis rhythm and a circadian degradation rhythm. The circadian synthesis rhythm was controlled mainly at the transcriptional level (Chen et al., 1996). For the circadian degradation rhythm, some factors were responsible for COP23 degradation. The addition of chloramphenicol to the cultures prevented COP23 degradation, so new protein(s) synthesis before the decline of COP23 must be essential for COP23 degradation. When EGTA was added to the Synechococcus RF-1 cultures before COP23 degradation, the decline of COP23 was prevented. Thus, extracellular Ca2+ was required for the circadian degradation of COP23. However, the addition of EGTA did not affect the synthesis rhythm of COP23. Light was also needed for COP23 degradation. Comparing the light spectra, blue light is more effective for COP23 degradation than white or red light. Phosphorylation could not be detected in vivo or in vitro before COP23 degradation. Thus, COP23 degradation seemed not to be attributed to its phosphorylation.
Keywords: Calcium requirement; Circadian degradation; Circadian rhythm; Light effect; Output pathway; Synechococcus RF-1.
Introduction
Circadian rhythms are a widespread phenomenon regulated by biological clocks. They are responsible for the daily rhythmicity found in a wide variety of organisms, including mammals, insects, higher plants, fungi, algae, and cyanobacteria. The circadian system is generally composed of three basic divisions: (a) a circadian oscillator; (b) input pathways of signal transduction for clock setting; and (c) output pathways of signal transduction for the temporal regulation of specific biological processes. Since a great deal of studies have focused on understanding the nature of the "oscillator," remarkable progress has been made regarding its components and regulation (Young, 1998; Dunlap, 1999; Johnson and Golden, 1999; Lowrey et al., 2000). Thus, the molecular mechanisms of "clock oscillators" have been widely studied in humans (Tei et al., 1997; Sun et al., 1997), mice (Antoch et al, 1997; King et al., 1997), Drosophila (Sehgal et al., 1995; Rosbash et al., 1996; Young et al., 1996), Neurospora (Dunlap, 1996; Crosthwaite et al., 1997), and cyanobacteria (Ishiura et al., 1998; Johnson and Golden, 1999).
In unveiling the circadian clock, it is important to identify how environmental cues such as light or temperature entrain the oscillator (input pathway) and how the information from the clock is transduced to regulate rhythmic cellular processes (output pathway) in addition to the regulation mechanism of the oscillator itself. Several research
works have studied input and output pathways (Huang and Grobbelaar, 1995; Liu et al., 1996; Tsinoremas et al., 1996; Anderson and Kay, 1997; Katayama et al., 1999; Kreps et al., 2000). Some of them were investigated at a biochemical level. For example, the effect of different light conditions on the circadian expression of the gene encoding chlorophyll a/b binding protein was studied for the input pathway (Somers et al., 1998; Bognar et al., 1999). In the control mechanism of the output pathway, the rhythm of CO2 metabolism in Bryophyllum (Wilkins, 1992) and that of bioluminescence or nitrate reductase in Gonyaulax (Morse et al., 1989; Morse et al., 1990; Ramalho et al., 1995) were examined. In addition, a molecular mechanism that links the feedback loop of the central circadian oscillator in the suprachiasmatic nucleus in mice to the regulation of a clock-controlled gene has been discovered (Jin et al., 1999).
In our previous studies, a 23 kDa circadian oscillating protein, COP23, located in the cell membrane of Synechococcus RF-1 was cloned and characterized (Chen et al., 1996). A search of the GenBank uncovered no homologous genes. Detection of COP23 in three other cyanobacteria Synechococcus PCC7942, Synechocystis PCC6803, and Cyanothece ATCC51142 by Southern hybridization using COP23 gene as a probe indicated that hybridization bands could only be detected in the genomic DNA of Cyanothece ATCC51142. Both Synechococcus RF-1 and Cyanothece ATCC51142 are nitrogen-fixing unicellular cyanobacteria. Thus, COP23 might exist exclusively in unicellular cyanobacteria of nitrogen fixation.
The exposure of Synechococcus RF-1 to a diurnal light/dark regimen induced not only a circadian synthesis
*Corresponding author. Fax: 886-2-2782-7954; E-mail:tanchi@gate.sinica.edu.tw