Bot. Bull. Acad. Sin. (2003) 44: 159-166
Wang et al. Spatial pattern of Cryptocarya chinensis
Spatial pattern of Cryptocarya chinensis life stages in lower subtropical forest, China
Zheng-Feng Wang1,*, Shao-Lin Peng1, Shi-Zhong Liu1, and Zhen Li2
1South China Institute of Botany, Chinese Academy of Sciences, Guangzhou 510650, P. R. China
2School of Life Sciences, Zhongshan University, Guangzhou 510275, P. R. China
(Received April 2, 2002; Accepted December 3, 2002)
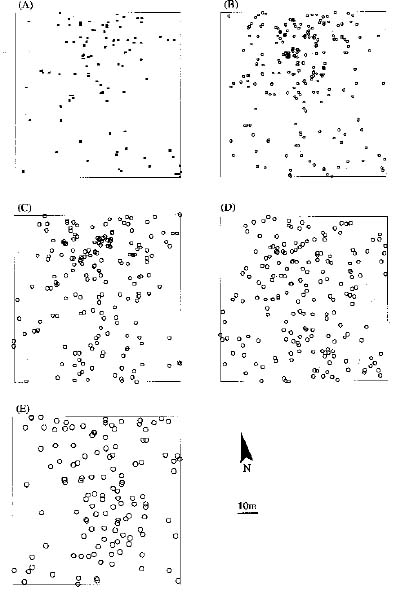
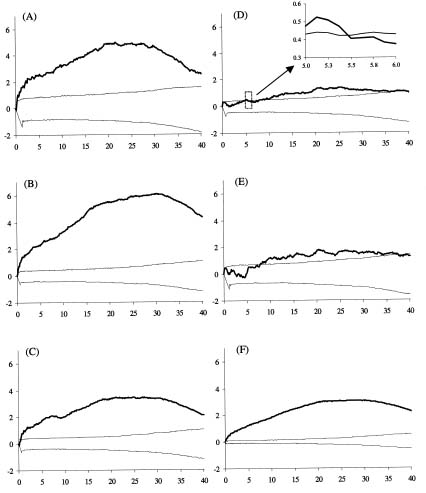
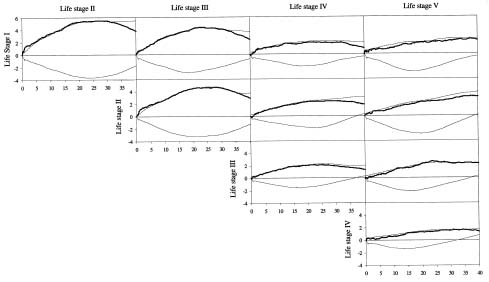
Abstract. Two aspects of life stages are important determinants in the spatial pattern of plant populations: one is spatial distribution of individual plants within life stages; the other is spatial association between different life stages. Using Ripley's univariate L(t) and bivariate L12(t) functions, we analyzed the spatial pattern within and between life stages of Cryptocarya chinensis in an 80 m × 80 m plot in lower subtropical China. Based on stem diameter and height, five life stages (I, II, III, IV, V) were distinguished. Ripley's univariate L(t) function showed clumped distributions within all life stages, with fine-scale random distributions in old life stages VI and V. From the bivariate L12(t) function, young life stages I, II and III appeared significantly attractive to each other at local and intermediate distances, but independently distributed regarding old life stages VI, V at almost all distances. The oldest life stage, V, was independently distributed relative to other life stages. Considering these results, we inferred that the existence of suitable survival environments (e.g. canopy gaps) are a determinant in shaping the spatial pattern of young life stages of C. chinensis; with increasing body size with growth, intraspecific competition develops, and a fine-scale random distribution appears within old life stages.
Keywords: Cryptocarya chinensis; Intraspecific competition; Life stage; Spatial pattern; Spatial statistics.
Introduction
Plant spatial patterns have long been a major issue in ecology (Ashby, 1935; Greig-Smith, 1983), and spatial patterns at various life stages or age classes have received considerable attention in ecological research (e.g. Lu, 1986; Sterner et al., 1986; Rebertus et al., 1989; Liu, 1990; Liang, 1992; Moeur, 1993; Xu and Zheng, 1993; Cai and Song, 1997; Barot et al., 1999; He and Li, 1999; Nanami et al., 1999; Dovciak et al., 2001). Since adults have undergone the life stages of seedlings, saplings and juveniles, an analysis of the spatial patterns of these early life stages may help reveal processes determining the spatial patterns at later life stages.
To do so, two aspects in life stage spatial pattern have to be analyzed. One is spatial distribution of individuals within life stages, and the other is spatial association between different life stages. Both of them represent important aspects of species' spatial pattern. Within life stages, thinning processes might increase distances between individuals of different cohorts and lead to regular distributions (Antonovics and Levin, 1980; Sterner et al., 1986; Kenkel, 1988; Moeur, 1993); On the other hand, offspring might be associated with some "nurse plants" (Callaway and Walker, 1997; Haase, 2001) or escape their conspecific adults due to more predation/pathogen in the periphery
of adults (Smith and Goodman, 1987; also see Barot et al., 1999), leading either to positive or negative associations between life stages. Environmental heterogeneity, seed dispersal and other factors also influence spatial patterns (Duncan, 1991; Barot et al., 1999; Dovciak et al., 2001).
Moreover, numerous studies have demonstrated that an individual plant competes for nutrient resources with its immediate neighbors (Mack and Harper, 1977; Antonovics and Levin, 1980; Connell et al., 1984; Moeur, 1993; Barot et al., 1999), leading to changes in the spatial patterns within and between life stages on both a small and large scale. Therefore, analysis of spatial pattern within and between life stages at different spatial ranges will aid in interpreting the species spatial and temporal dynamics during species population development.
In lower subtropical China there have been no reports to date of plant spatial pattern in different life stages at different distance ranges. Here we present our research on Cryptocarya chinensis (Hance) Hemsl. (Lauranceae) in this region. Our aim was to (1) detect the dynamic aspects of spatial pattern at different life stages; (2) examine the spatial association between life stages; and to (3) discuss the possible mechanisms shaping the species spatial pattern. As one project to study the role of the spatial dynamics in determining species response to forest fragmentation on time scales of generation, our study, using C. chinensis in continuous lower subtropical monsoon evergreen broad-leaved forest, will also provide baseline information on the natural dynamics of C. chinensis.
*Corresponding author. Tel: +86-20-85232862; Fax: +86-20-85232615; E-mail: wzf@scib.ac.cn