Bot. Bull. Acad. Sin. (2003) 44: 179-186
Hyodo et al. Ethylene biosynthesis in infected tissue
Ethylene biosynthesis in sweet-potato root tissue induced by infection with black rot fungus (Ceratocystis fimbriata)
Hiroshi Hyodo*, Seijiro Yoshioka, Yoshifumi Imai, Hiroko Nakane, and Fumie Nishikawa
Department of Biological Sciences, Faculty of Agriculture, Shizuoka University, Ohya, Shizuoka 422-8529, Japan
(Received December 19, 2002; Accepted February 24, 2003)
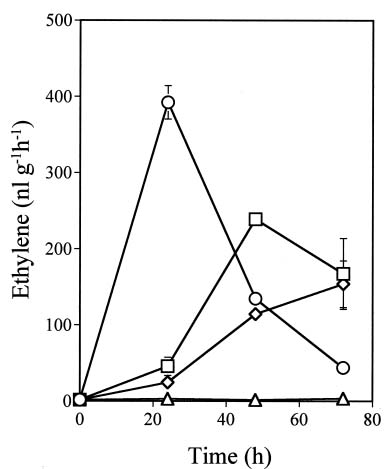
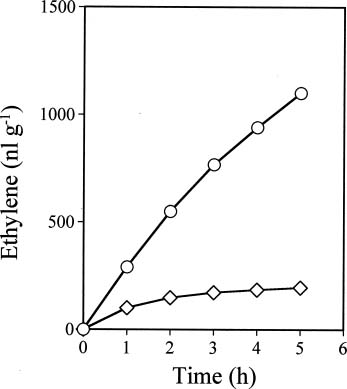
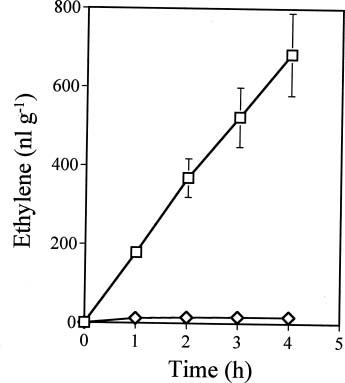
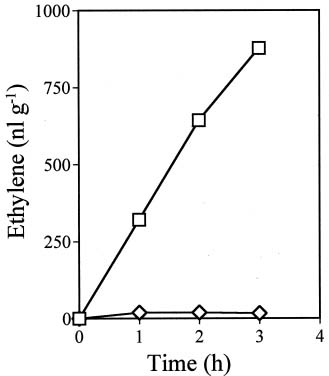
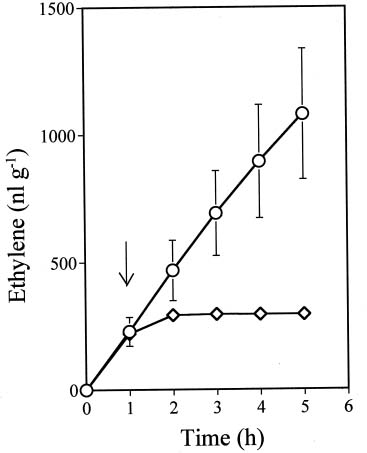
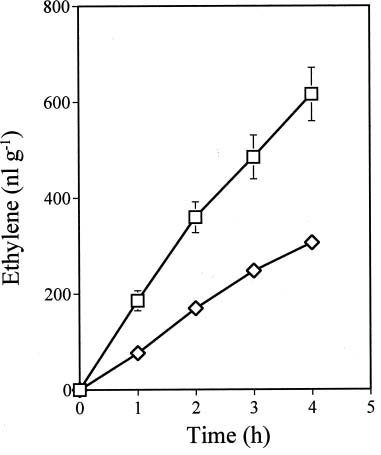
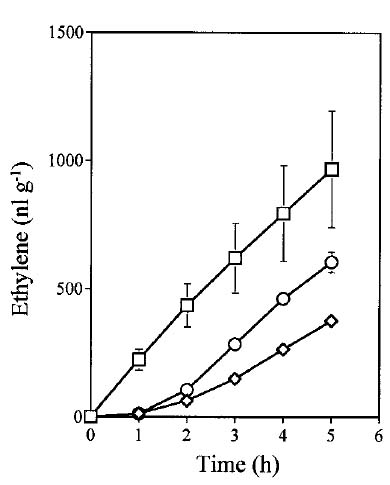
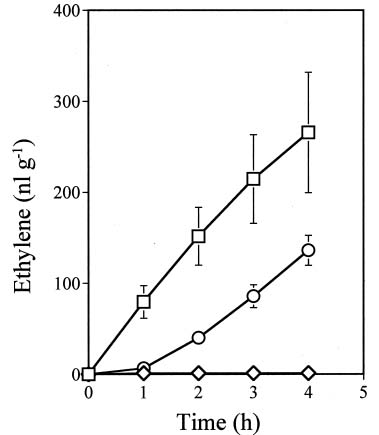
Abstract. Ethylene production rapidly increased in sweet potato (Ipomoea batatas Lam.) root tissue in response to infection by black rot fungus (Ceratocystis fimbriata Ell. & Halst). Ethylene production increased in the surface layer (0-0.5 mm) of the root inoculated with endoconidia of C. fimbriata, reaching a maximum 1 day after inoculation and declining thereafter. The rise in the surface layer was followed by a subsequent increase in the inner, second (0.5-1.0 mm), and third (1.0-1.5 mm) layers as the fungus penetration progressed. Discs 10 mm in diameter and 0.5 mm thick excised from the surface of the infected root slices were used for the experiments. Our previous study demonstrated that ethylene synthesis in the infected tissue may occur via a pathway independent of the methionine-ACC (1-aminocyclopropane-1-carboxylic acid) pathway. Ethylene synthesis in the infected tissue was significantly suppressed by the inhibitors that interfere with NADPH oxidase, phospholipase A2 and lipoxygenase, by metal ion chelators, and by scavengers of reactive oxygen species. The inhibition of metal ions was restored by the addition of cupric ion. These results suggest that in the infected sweet potato root tissue, polyunsaturated fatty acids are released from membrane phospholipids followed by peroxidation by lipoxygenase. The hydroperoxides could be decomposed in the presence of copper ion or copper enzyme and reactive oxygen species such as hydroxyl radical, leading to ethylene generation. De novo protein synthesis was required for ethylene synthesis to be induced in response to the fungal infection.
Keywords: Ceratocystis fimbriata; Copper ion; Ethylene biosynthesis; Infected tissue; Ipomoea batatas; Lipoxygenase; Membrane lipids; Reactive oxygen species; Unsaturated fatty acids.
Introduction
When sweet potato (Ipomoea batatas Lam.) roots are infected by black rot fungus (Ceratocystis fimbriata Ell. & Halst), a wide range of metabolic changes is induced in the host tissue (Uritani, 1971). These changes include an increase in respiration and stimulation of polyphenol synthesis, coumarin synthesis, furanoterpenoid synthesis, and ethylene synthesis (Uritani, 1967; 1971; Kojima, 1999). It has been reported that a large amount of ethylene is produced in response to infection with C. fimbriata in sweet potato roots (Stahmann et al., 1966; Imaseki et al., 1968a; Sakai et al., 1970; Kato and Uritani, 1972; Hyodo and Uritani, 1984; Hirano et al., 1991; Okumura et al., 1999).
The plant hormone ethylene plays an essential role in the growth, development, and senescence of higher plants and is synthesized normally by the pathway mediated by methionine and ACC (1-aminocyclopropane-1-carboxylic acid) (Adams and Yang, 1979; Yang and Hoffman, 1984). In the methione-ACC pathway, ACC synthase and ACC oxidase have a pivotal role in regulating ethylene
synthesis. In lower plants, however, it is known that ACC fails to serve as a direct intermediate in ethylene biosynthesis (Osborne et al., 1996). Ethylene is also produced as a result of rapid response to abiotic and biotic stresses such as wounding and disease in plants (Yang and Hoffman, 1984; Boller, 1991; Hyodo, 1991; Abeles et al., 1992). In diseased tissue ethylene production is caused by host-parasite interactions.
In sweet potato roots it has been considered that most of the ethylene is generated in the living host cells adjacent to the necrotic region invaded by the fungus (Hirano et al., 1991; Okumura et al., 1999; Hyodo et al., 2003). A previous study has suggested that major sources of ethylene produced in sweet potato root tissue after infection by the fungus are derived from a pathway distinct from the methionine-ACC or glutamate pathway (Hyodo and Uritani, 1984; Hirano et al., 1991; Okumura et al., 1999; Hyodo et al., 2003).
The aim of the present study was to investigate the mechanism by which ethylene is biosynthesized in sweet potato root tissue infected by C. fimbriata. Our results suggest that phospholipase, lipoxygenase, reactive oxygen species, and copper ion are involved in ethylene formation.
*Corresponding author. Tel: +81-54-238-4830; Fax: +81-54-237-3028; E-mail: abhhyou@agr.shizuoka.ac.jp