Bot. Bull. Acad. Sin. (2003) 44: 193-198
Li et al. Expression of TDC in chloroplasts of tobacco
Expression of tryptophan decarboxylase in chloroplasts of transgenic tobacco plants
Qiu-Rong Li2,3, Stefano Di Fiore3, Rainer Fischer3, and Miao Wang1,*
1Institute of Applied Ecology, The Chinese Academy of Sciences, Shenyang 110016, The People's Republic of China
2General Hospital in Nanjing Millitary District, Nanjing 210002, The People's Republic of China
3Botanical Institute of Rheinisch-Westfalisch Technische Hochschule Aachen, D-52074 Aachen, Germany
(Received August 5, 2002; Accepted February 17, 2003)
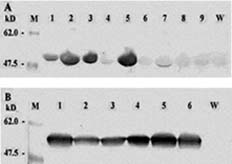
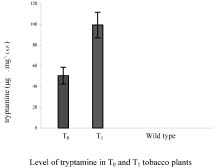
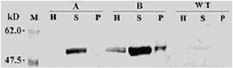
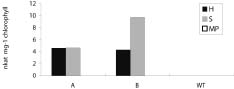
Abstract. Tryptophan decarboxylase (TDC) is a key enzyme in the biosynthesis of terpenoid indole alkaloids (TIAs) from plants such as Catharanthus roseus. We transformed tobacco plants with tdc gene and expressed chloroplast-targeted versions of TDC protein. TDC was shown to be over-expressed in chloroplasts of tobacco plants by Western blot analysis and TDC enzymatic assay. The effective expression of TDC lead to accumulation of tryptamine in transgenic tobacco plants. T1 plants showed higher levels of TDC expression than T0 plants. In order to confirm that TDC was correctly targeted and expressed in chloroplasts, they were isolated from T1 transgenic tobacco plants, and the different fractions of purified chloroplasts were analyzed. Transgenic plants displayed the phenotype characteristic of necrotic syndromes, which was related to the accumulation of tryptamine.
Keywords: Chloroplast; Nicotiana tabacum; Terpenoid indole alkaloids; Tryptophan decarboxylase.
Introduction
Tryptophan decarboxylase (TDC, EC 4.1.1.28) is the key enzyme in the early step of the TIA biosynthetic pathway and links primary metabolism to secondary metabolism by converting tryptophan into tryptamine. Tryptamine is subsequently condensed with secologanin, resulting in strictosidine, the precursor to pharmaceutically important alkaloids such as vinblastine and vincristine (Hashimoto and Yamada, 1994). Catharanthus roseus (Madagascar periwinkle, the family of Apocynaceae) produces these important alkaloids (Meijer et al., 1993). Vinblastine and vincristine are mainly extracted from C. roseus, but there are only trace amount of the alkaloids. Cell cultures of C. roseus had been considered to be sources of medicinally important TIAs, but they suffered from low productivity (Pasquali et al., 1992; Roewer et al., 1992; Moreno et al., 1995). TDC enzymatic activity is one of the bottlenecks in the TIA biosynthetic pathway. A cDNA clone encoding TDC was first isolated from C. roseus (De Luca et al., 1989). A high accumulation level of tryptamine was observed by expressing TDC in transgenic tobacco plants (Songstad et al., 1990). However, there was no report found in targeting and expressing recombinant TDC enzymes in subcellular compartments to study TIA biosynthesis. TDC is a cytosolic soluble enzyme in TIA-producing plants such as C. roseus. Tobacco is an ideal host plant, and endogenous TDC activity is not detected in it because it does not contain tdc gene in its genome or the enzymes of
downstream to metabolize tryptamine, so the biosynthesis of tryptamine in the leaves of transgenic tobacco plants expressing targeted TDC was used as a direct evidence of in vivo TDC function in the chloroplasts. The chloroplast represented an important subcellular compartment because the chloroplast is the site of L-Tryptophan biosynthesis. In this paper, we manipulated the reaction catalyzed by TDC to increase the level of tryptamine through engineering the TIA biosynthetic pathway in plants. TDC was expressed in the chloroplasts of transgenic tobacco plants to study the effect of targeting TDC to chloroplast on the in vivo functionality in tobacco. The special phenotypes of transgenic plants expressing recombinant TDC in the chloroplast were observed.
Materials and Methods
Plant: Nicotiana tabacum L. cv. Petite Havana SR1.
Bacteria: Agrobacterium tumefaciens GV3101 (pMP90RK, GmR, KmR, Rif R) (Koncz and Schell, 1986).
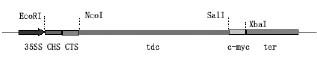
Manipulation of DNA was performed by standard techniques (Sambrook et al., 1986). Tdc was amplified by PCR using the pTSK plasmid DNA (Leech et al., 1998) with the sense primer 5´-CGCGAGCTCCATGGGCAGCATTGATTCAAC-3´ and the antisense primer 5´-CCCAAGCTTGTCGACGGCTTCTTTGAGCAAATCATC-3´ . These primers were designed to amplify tdc downstream to the 5´ untranslated region (UTR) and upstream to the stop codon of the cDNA sequence (Genebank accession number M25151) in the pTSK plasmid DNA. 5´ SacI and NcoI
*Corresponding author. Tel: 0086-024-23051460; Fax: 0086-024-23843313; E-mail: wangmiaojue@163.com