Bot. Bull. Acad. Sin. (2003) 44: 199-210
Lin et al. cry1C in plastid confers high insecticidal efficacy
Expression of a Bacillus thuringiensis cry1C gene in plastid confers high insecticidal efficacy against tobacco cutworm - a Spodoptera insect
Chi-Hui Lin1,4, Yen-Yu Chen1, Ching-Chou Tzeng2, Hsin-Sheng Tsay3,4, and Liang-Jwu Chen1,*
1Institute of Molecular Biology, National Chung Hsing University, Taichung, Taiwan 402
2Department of Biopesticide, Taiwan Agricultural Chemicals and Toxic Substances Research Institute, Wufeng, Taichung, Taiwan 413
3Department of Agronomy, Taiwan Agricultural Research Institute, Wufeng, Taichung, Taiwan 413
(Received January 7, 2003; Accepted April 11, 2003)
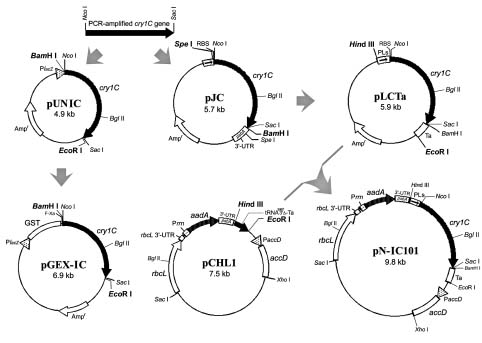
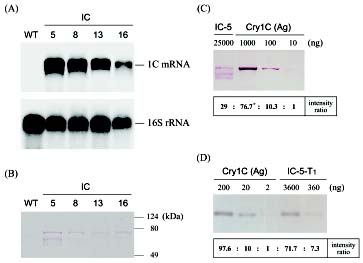
Abstract. We have demonstrated that the toxic domain of a wild-type Bacillus thuringiensis (Bt) cry1C gene could be expressed efficiently in the chloroplasts of transplastomic tobacco plants, resulting in high resistance to tobacco insects. A DNA fragment with the toxic domain of a wild-type Bt cry1C gene was constructed under the control of rbcL promoter and an E. coli thra terminator in a plasmid vector flanked by chloroplast sequences. This plasmid vector was then delivered into the chloroplast through biolistic bombardment technique. Plant transformants that possessed spectinomycin and streptomycin resistance were selected, and the integration of cry1C gene in these transformants was confirmed by PCR and Southern analyses. Results from PCR and Southern hybridization indicated that the truncated cry1C gene was integrated into the chloroplast genome as we expected through homologous recombination. High expression level of the integrated Bt cry1C gene in the chloroplasts was observed through Northern and Western hybridization assays. These transplastomic tobacco plants are highly toxic to the tobacco cutworm Spodoptera litura, causing a mortality rate of 76.9% to 100% after 72 h of feeding. The truncated Bt toxin is expressed in high levels in the chloroplast (about 1% of total proteins) and the plastid transgenes were known not to transmit through pollen. So that, this study will facilitate not only improvement in breeding for insect-resistant plants, but also the prevention of contamination of transgenes among crop plants.
Keywords: Bacillus thuringiensis; Chloroplast; cry1C gene; Insecticidal protein; Nicotiana tabaccum; Plastid transformation; Spodoptera lituar.
Abbreviations: BA, N6-benzyl adenine; BAW, beet armyworm; Bt, Bacillus thuringiensis; FAW, fall armyworm; GST, glutathione S-transferase; ICP, insecticidal crystal protein; MCCW, Mediterranean climbing cutworm; NAA, a-naphthalene acetic acid; PCR, polymerase chain reaction; RBS, ribosomal binding site; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; TCW, tobacco cutworm.
Introduction
Crystal (Cry) proteins from Bacillus thuringiensis (Bt) have an insecticidal effect on a number of insects. Most Bt toxins are synthesized as protoxins that are then proteolytically converted into active toxin fragments (ca. 60-70 kDa) in the insect midgut. The activated toxin interacts with the midgut epithelium cells of susceptible insects and generates pores in the cell membranes, disturbing the osmotic balance of cells. This causes the insect larvae to stop feeding and eventually die (Gill et al., 1992). Attempts to express Bt toxin genes in plants through nuclear genome transformation have been very successful (Barton
et al., 1987; Fischhoff, 1987; Vaeck et al., 1987; Perlak et al., 1990; Cheng et al., 1992; Fujimoto et al., 1993; Koziel et al., 1993; Van der Salm et al., 1994; Stewart et al., 1996; Nayak et al., 1997; Cheng et al., 1998). However, the expression levels of original unmodified Bt toxin genes in plants were extremely low, in the range of 0.001 - 0.005% of total soluble proteins (Barton et al., 1987; Fischhoff, 1987; Vaeck et al., 1987). Numerous factors have been proposed as the cause of this low expression level, such as premature transcription termination, aberrant mRNA splicing, mRNA instability, and inefficient codon usage (Perlak et al., 1991). The synthetic Bt toxin genes designed to avoid many of the undesirable features and unfavorable expression in plants have resulted in dramatic increases of Bt gene expression levels, up to 0.1-0.3% of total soluble proteins (Perlak et al., 1990; Fujimoto et al., 1993; Koziel et al., 1993; Van der Salm et al., 1994; Cheng et al., 1998). However, artificial synthesis of large Bt toxin genes is laborious and cost inefficient and, therefore, has limited applications in the long run.
*Corresponding author. Tel: +886-4-22851885; Fax: +886-4-22874879; E-mail: ljchen@dragon.nchu.edu.tw
4Present address: C-H Lin: Institute of Chemistry, Academia Sinica, Taipei, Taiwan. E-mail: chlin824@giga.net.tw; H-S Tsay: Department of Applied Chemistry, Chaoyang University of Technology, Wufeng, Taichung, Taiwan 413.