Bot. Bull. Acad. Sin. (2003) 44: 223-228
Hao et al. Phylogenetics of Bauhinia subgen. Phanera
Phylogenetics of Bauhinia subgenus Phanera (Leguminosae: Caesalpinioideae) based on ITS sequences of nuclear ribosomal DNA
Gang Hao, Dian-Xiang Zhang*, Ming-Yong Zhang, Li-Xiu Guo, and Shi-Jin Li
South China Institute of Botany, The Chinese Academy of Sciences, Guangzhou 510650, P. R. China
(Received October 2, 2002; Accepted April 11, 2003)
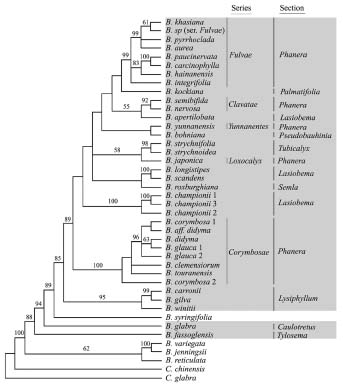
Abstract. The pantropical legume genus Bauhinia is currently subdivided into four subgenera. Subgenus Phanera, with ca. 150 species, is the largest. A parsimony analysis was conducted on ITS sequence data from 38 species, with representatives from all four subgenera, and species sampled from nine of 11 sections of subgen. Phanera, and including two Cercis species as the outgroup. The well-resolved strict consensus tree suggests that subgen. Phanera and the monotypic subgen. Barklya together form a monophyletic group. In the Phanera-Barklya clade, the African (sect. Tylosema), American (sect. Caulotretus), and Australasian (subgen. Barklya, subgen. Phanera sect. Lysiphyllum) species are the first (paraphyletic) lineages, while the Asian taxa (except species of sect. Lysiphyllum) form a monophyletic group. The ITS tree also revealed that sects. Phanera and Lasiobema are not monophyletic. Monophyletic groups supported by the phylogenetic analysis include ser. Fulvae and ser. Corymbosae of sect. Phanera, sect. Tubicalyx, and the Asian-Australasian sect. Lysiphyllum.
Keywords: Bauhinia; Caesalpinioideae; ITS; Leguminosae; Phylogenetic analysis; subgen. Phanera.
Introduction
Bauhinia L., a pantropical genus of about 300 species (Wunderlin et al., 1987) of trees, shrubs or lianas with bifoliolate, bilobed or simple entire leaves, has always been controversial in its taxonomy, including whether or how it could be divided into small genera.
According to Wunderlin et al. (1981, 1987), the ancestor of the genus gave rise to two main phyletic lines, one to the mostly arborescent or shrubby subgenera Bauhinia, Elayuna and Barklya, the second to subgenus Phanera, which mostly comprises lianas. A morphological cladistic analysis of the genus by Zhang (1995) and a survey of the rpL2 chloroplast intron for 78 species by Lai et al. (1997), however, both yielded results not fully compatible with the conclusions of Wunderlin et al.
Subgenus Phanera, comprising about 150 species, is the largest subgenus in Bauhinia. Pantropical in distribution, but with most of its species occurring in continental Asia and the Malesia region, the subgenus is subdivided into 11 sections (Wunderlin et al., 1987). Several of these sections are further subdivided into subsections or series.
In their recent phylogenetic analysis of the subfamily Caesalpinioideae based on chloroplast trnL intron sequences, Bruneau et al. (2001) suggested that the sub
genus Phanera, as well as the whole genus Bauhinia, might not be monophyletic.
Although taxonomic and floristic studies on the genus Bauhinia in general, and on subgen. Phanera in particular, have helped improve our knowledge of the group (Chen, 1988; Larsen and Larsen, 1973, 1983, 1991, 1996; Larsen et al., 1980, 1984; Zhang, 1993), systematic studies on the genus are still hampered by its large number of species, prevalence of morphological convergence, and difficulties in obtaining material. In this study, we newly sequenced nuclear ITS regions from representative species of Bauhina, with the aim of assessing the monophyly of subgen. Phanera as currently defined, and of clarifying the intra-subgeneric relationships within the group.
Materials and Methods
Taxon Sampling
A total of 38 species were included in this molecular study (Table 1). For the ingroup we sampled 40 accessions representing 32 species of subgen. Phanera, as well as two species of subgen. Bauhinia, one species of the monotypic subgen. Barklya, and one species of subgen. Elayuna. The ingroup taxa chosen covered all four subgenera recognized by Wunderlin et al. (1987). The 32 species of subgen. Phanera represented nine of the 11 sections (Wunderlin et al., 1987). Most of the species chosen were from the two larger sections that are apparently polymorphic, viz., sect. Lasiobema and sect. Phanera. For some sections, e.g., sects. Palmatifolia, Tubicalyx, and
*Corresponding author. Tel: 86-20-8523-2043; Fax: 86-20-8523- 2831; E-mail: dx-zhang@scib.ac.cn