Bot. Bull. Acad. Sin. (2003) 44: 229-238
Liao et al. Regeneration of coniferous-broadleaf forest in YYL
Regeneration patterns of yellow cypress on down logs in mixed coniferous-broadleaf forest of Yuanyang Lake Nature Preserve, Taiwan
Chi-Cheng Liao1, Chang-Hung Chou2, and Jiunn-Tzong Wu1,3,*
1Department of Botany, National Taiwan University, Taipei 106, Taiwan
2Graduate Institute of Tropical Agriculture, National Pingtung University of Science and Technology, Pintung 912, Taiwan
3Institute of Botany, Academia Sinica, Taipei 115, Taiwan
(Received August 26, 2002; Accepted May 6, 2003)
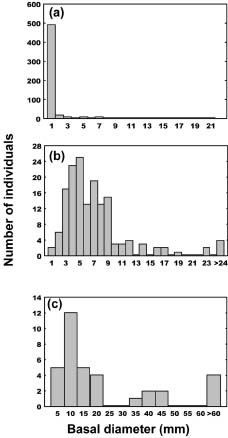
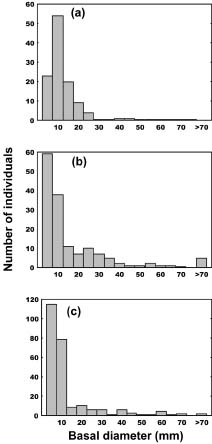
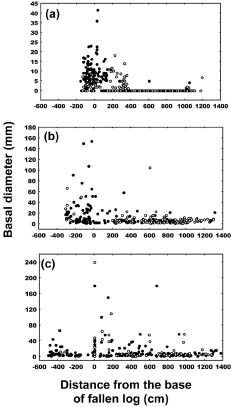
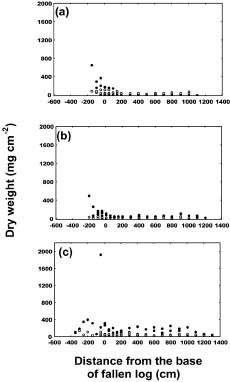
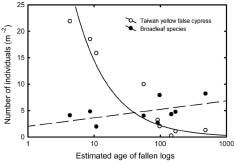
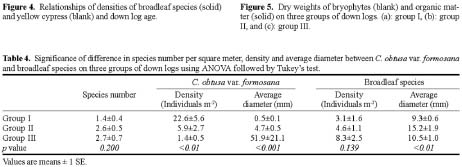
Abstract. The importance of down logs in forest gaps for forest development, tree regeneration, and the co-existence of yellow cypress (Chamaecyparis obtusa Sieb. & Zucc. var. formosana (Hayata) Rehder) and broadleaf trees was studied in the temperate forest of Yuanyang Lake Nature Reserve (YYL), Taiwan. On nine down logs in a plot of 0.25-ha, seedlings with diameter at breast height larger than 1 cm were recorded. It was found that the species occurring on the sampled down logs were the same as those in the studied area. Few broadleaf trees could reach the height of 10 m necessary to change the current canopy layer, which mainly consisted of yellow cypress. Regeneration of trees on down logs was sufficient to maintain the forest community, including its floristic composition, canopy structure, and species diversity. Tree seedlings of different species also varied in their distribution patterns on down logs. Yellow cypress seeds germinated on new down logs, but not on root swellings. The seedlings grew to fill gaps, while density was reduced from young to old down logs, possibly due to intra-species competition. Epiphytic bryophytes on new down logs seemed to provide a suitable habitat for the germination of yellow cypress seeds. In contrast, broadleaf trees grew mainly on the base and root swellings of new down logs and gradually recruited their seedlings onto other parts of older down logs. Availability of nutrient resources seemed to be the important determinant for the competition between seedlings of cypress and broadleaf plants. The niche partition hypothesis was employed to explain the regeneration of cypress plants in this nature preserve.
Keywords: Chamaecyparis obtusa var. formosana; Down log; Niche partitioning; Regeneration; Yellow cypress; Yuanyang Lake Nature Preserve.
Introduction
Gap dynamics, including tree death, seedling regeneration, and adult replacement, have been studied in temperate (Runkle, 1981; Runkle, 1982; Veblen, 1986; Yamamoto, 1988; Nakashizuka, 1989; Lertzman, 1992; Gray and Spies, 1996; Runkle, 2000) and tropical forests (Brokaw, 1985; Arriaga, 1988; Lawton and Putz, 1988; Uhl et al., 1988; Dalling et al., 1998), but have received little attention in the temperate forests of Taiwan. Previous studies have considered succession trends of the forests (Chang, 1961; Liu, 1971), but not gap dynamics or regeneration of trees.
Chamaecyparis obtusa Sieb. & Zucc. var. formosana (Hayata) Rehder (yellow cypress) is one of the dominant tree species in the temperate forests in Taiwan (Su, 1984; Jen, 1995). Yellow cypress was reported to establish seedlings on down logs and mineral soils in some Taiwan for
ests (Liu et al., 1961), but no further studies were done on this phenomenon. Most studies concerning recruitment of yellow cypress has been done in plantations, and they concluded that thinning broadleaf trees could promote establishment of yellow cypress seedlings by reducing inter-specific competition (Hung, 1984; Chiu et al., 1995; Lo-Cho et al., 1999). However, this conclusion was based on the premise that the resource requirements of broadleaf and yellow cypress trees overlap and that yellow cypress is a weak competitor. There is some uncertainty regarding these viewpoints.
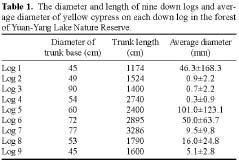
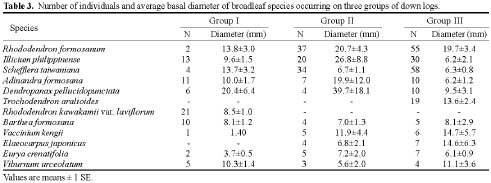
Yellow cypress and broadleaf trees are known to co-exist in a forest at Yuanyang Lake Natural Preserve (YYL), northern Taiwan (Chou et al., 2000). This virgin mixed coniferous-broadleaf forest is free from human disturbance, and it is of interest to consider the mechanisms that might allow the trees to coexist in this natural system. Based on past research, both broadleaf trees and yellow cypress seedlings occur in abundance on down logs (Liu et al., 1961). The role of these logs may therefore be important to the regeneration and co-existence of trees in YYL.
*Corresponding author. Tel: 886-2-2789-9590 ext 420; Fax: 886-2-2782-7954; E-mail: jtwu@gate.sinica.edu.tw