Bot. Bull. Acad. Sin. (2004) 45: 111-118
Chen et al. Purification and characterization of carboxymethyl cellulase
Purification and characterization of carboxymethyl cellulase from Sinorhizobium fredii
Po-Jui Chen1, Tao-Chun Wei2, Yao-Tsung Chang2, and Liang-Ping Lin2,*
1Department of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan, Republic of China
2Institute of Microbiology and Biochemistry, National Taiwan University, Taipei 106, Taiwan, Republic of China
(Received July 12, 2001; Accepted January 13, 2004)
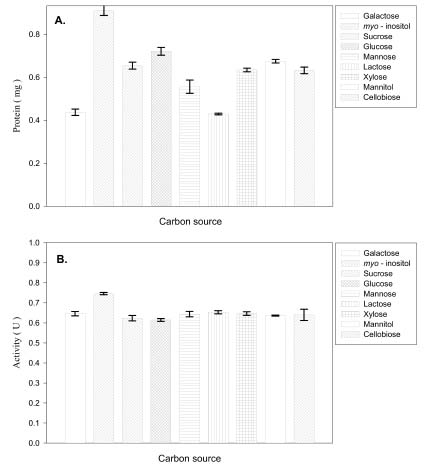
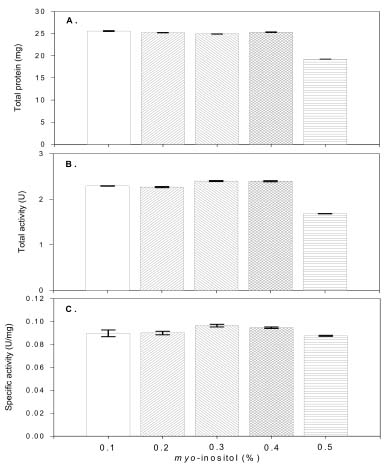
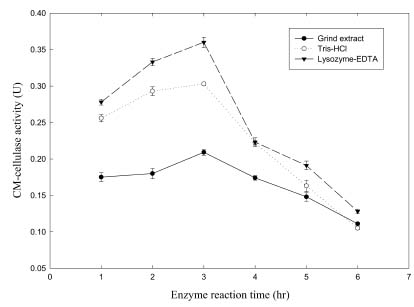
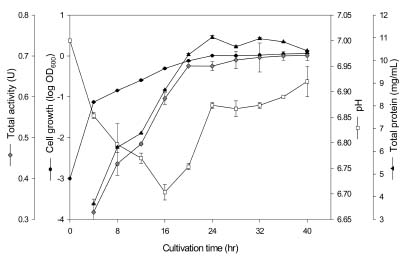
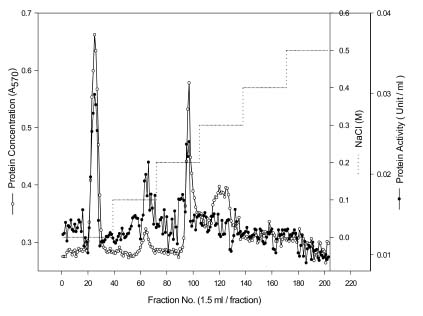
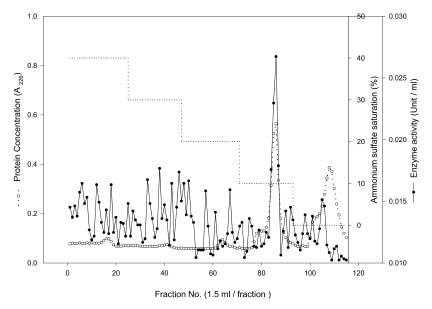
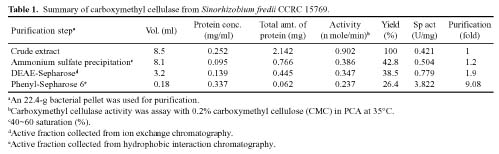
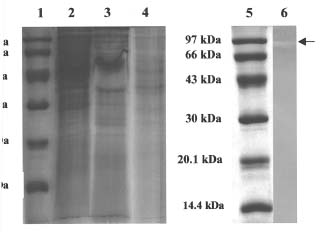
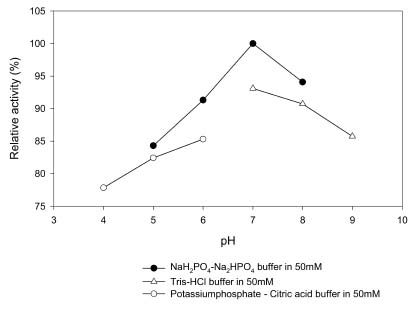
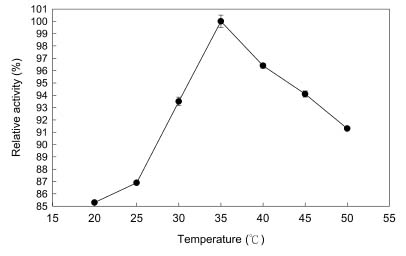
Abstract. Carboxymethyl cellulase (CMCase) is the key enzyme used by bacteria to decompose the plant root hair-wall during symbiosis. In this study, CMCase activities were routinely assayed using carboxymethyl cellulose (CMC) as a substrate. Coupled with the catalytic reaction of BCA (2,2'-bicinchoninic acid) solution, the color developed at 570 nm was measured to determine the enzyme activity. Sinorhizobium fredii CCRC 15769 was utilized to produce CMCase for this study. The optimum growth conditions of the cultural medium were studied in order to obtain sufficient cells for further CMCase purification, and further, for large-scale fermentation production. BIII medium containing 0.3% myo-inositol proved to be a suitable carbon source for bacterial cell growth and specific activity of CMCase. A 22.4 g wet weight of cells can be harvested from 3.5 liters of fermented medium through continuous centrifugation. The cells were suspended in 50 mM potassium phosphate-citric acid (PCA) buffer at pH 5.2 and disintegrated through ultra-sonication to obtain crude extract. Then, certain precipitates were collected at 40-60% ammonium sulfate saturation via ammonium sulfate fractionation of the crude extract. The supernatant obtained following centrifugation was loaded onto a DEAE Sepharose anion-exchange column, and the active fractions were collected and dialyzed against 10 mM Tris-HCl buffered at pH 7.4. For further purification, the dialyzed fraction was loaded onto a Phenyl-Sepharose column. The active fractions were dialyzed and then assayed by SDS-PAGE activity stain to confirm that it contained CMCase activity at the fraction of 94 kDa. The characterizations of CMCase were demonstrated as follows: the optimal temperature and pH were 35°C and 7.0, respectively. The purification fold was 9.08, and the recovery yield was 26.4%, and the specific activity was 3.822 U mg-1.
Keywords: Carboxymethyl cellulase; Sinorhizobium fredii; Protein extracts.
Abbreviations: CMCase, carboxymethyl cellulase; BCA, 2,2'-bicinchoninic acid; PCA, potassium phosphate-citric acid.
Introduction
Rhizobia are gram-negative bacteria that can induce morphologically distinct nodules on leguminous plant roots. The symbiotic interaction results in the inclusion of highly differentiated bacterial cells in host plant nodule cells, where they reduce atmospheric dinitrogen to ammonia, which is then utilized by the host plant. Establishment of an intracellular symbiosis between rhizobia and plants, which produces dinitrogen fixation, requires mutual recognition and signaling, penetration of the host cell, and finally nodulation and survival of bacteria in bacteroids. Carboxymethyl cellulase is probably involved in all of the infection stages.
The precise mechanism by which Rhizobium infects temperate legumes successfully remains unknown. Nutman (1956) proposed that rhizobia invade root hairs via an invagination process of the plant cell wall, which implies that the bacteria do not penetrate the root cell. Ljunggren and
Fahraeus (1961) proposed that polygalacturonase production by plant root cells, induced by homologous strains of Rhizobium, results in an increase in plant cell wall softening at the infection site on the root hair, thus allowing the bacteria to penetrate the cell membrane and initiate an infection thread. Mateos et al. (1992) proposed that the enzyme from Rhizobium leguminosarum bv. trifolii degraded both carboxymethyl cellulose and polypectate model substrate. This produced a localized degradation that completely traversed the root hair wall and facilitated direct bacterial penetration.
Electron microscopic studies of the infection process confirm a localized degradation of the root hair wall, indicating involvement of cell wall hydrolytic enzymes in the penetration process (Higashi and Abe, 1980; Callaham and Torrey, 1981; Ridge and Rolfe, 1985; Turgeon and Bauer, 1985). We are concerned with the possibility that the wall-degradation enzymes involved in the process are either associated with the bacteria or are locally induced in the plant by bacterial components.
Infection between rhizobia and legume roots is a delicately balanced process (Dazzo and Hubbell, 1982). If the infected root hair did not remain intact and viable, the in
*Corresponding author. Tel: 886-2-23635523; Fax: 886-2-23626455; E-mail: m046@ccms.ntu.edu.tw