Bot. Bull. Acad. Sin. (2004) 45: 119-125
Yang et al. Allelopathic phenolics and chlorophyll accumulation
Effects of three allelopathic phenolics on chlorophyll accumulation of rice (Oryza sativa) seedlings: II. Stimulation of consumption-orientation
Chi-Ming Yang1,*, Ing-Feng Chang1, Shu-Jin Lin1, and Chang-Hung Chou2
1Institute of Botany and Research Center for Biodiversity, Academia Sinica, Nankang, Taipei, Taiwan 11529, Republic of China
2Institute of Tropical Agriculture and Research Center for Biodiversity and International Cooperation, National Pingtung University of Science and Technology, Neipu, Pingtung, Taiwan 912, Republic of China
(Received May 2, 2002; Accepted November 11, 2003)
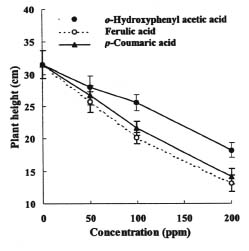
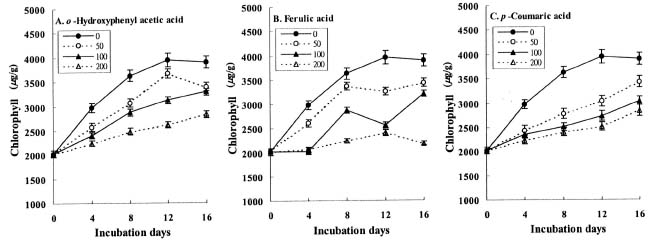
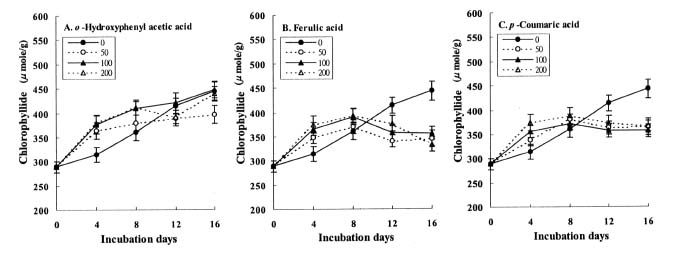
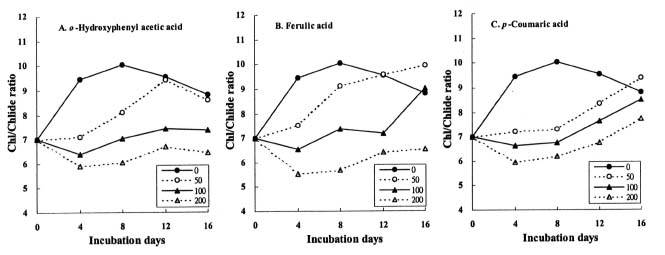
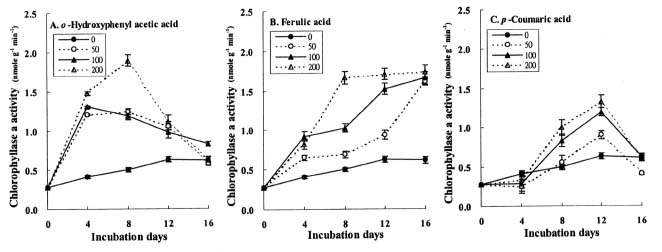
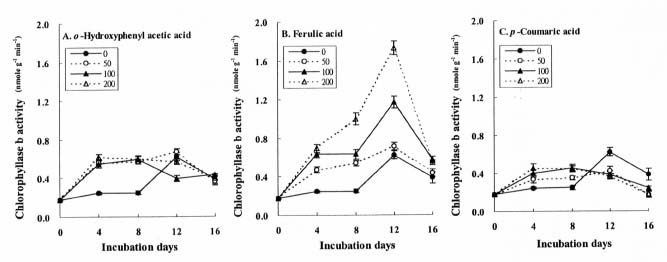
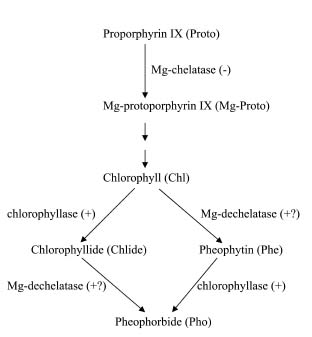
Abstract. The effects of three allelopathic phenolics, o-hydroxyphenyl acetic, ferulic and p-coumaric acids, on the chlorophyllase activity of rice leaf (Oryza sativa cv. TN67) were investigated. Ten-day-old green seedlings of rice were cultured in greenhouse for 16 d in Kimura's culture solution, which was changed every 4 days, with or without 50, 100 or 200 ppm of the phenolic compounds. Just before changing the culture solution, leaves were harvested to determine their chlorophyll (Chl) and chlorophyllide (Chlide) contents, and their chlorophyllase a and b activities. While the Chl and Chlide contents decreased and increased, respectively, causing the molar ratio of Chlide/Chl to increase, as the phenolic concentrations increased; the chlorophyllase a and b activities drastically increased. This suggests that the consumption-orientation of Chl was significantly stimulated by the exogenously applied phenolics. The order of inhibition of growth of the rice seedlings is: ferulic acid>p-coumaric acid>o-hydroxyphenylacetic acid. The order of inhibition effect on Chl accumulation is: p-coumaric acid>o-hydroxyphenylacetic>ferulic acid. The order of stimulation effect on chlorophyllase a activity is: o-hydroxyphenylacetic acid>ferulic acid>p-coumaric acid. The order of promotion effect on chlorophyllase b is: ferulic acid>o-hydroxyphenylacetic acid>p-coumaric acid. The different responses of chlorophyllase a and b activities to the same concentrations of allelochemical phenolics suggest that they may be two different enzymes. It is apparent that the three phenolics may enhance the activities of enzymes, such as chlorophyllase and Mg-dechelatase, responsible for the Chl degradative pathway. A combination of the present and the preceding data strongly suggest that the three allelopathic phenolics may comprehensively affect the biosynthetic and degradative pathways of Chl.
Keywords: Allelopathic phenolics; Chlorophyll; Chlorophyllase; Chlorophyllide; Consumption-orientation; Ferulic acid; Oryza sativa; o-hydroxyphenyl acetic acid; p-coumaric acid.
Introduction
More than 10,000 secondary metabolites in plants, such as phenolics, terpenoids, alkaloids, fatty acids, steroids, and polyacetylenes, are known to play an important role in allelopathy, which includes positive and negative effects (Inderjit, 1996; Olofsdotter, 1998; Rice, 1984; Waller, 1987). Even so, it is still largely a mystery how the allelochemicals, of the targeted organelle of the targeted cell of the targeted tissue of a targeted plant, function in the targeted reaction, during replication, transcription, translation, or post-translation (Kohli et al., 1998).
The multiple effects resulting from allelopathic phenolics include decreases in plant growth, absorption of water and mineral nutrients, ion uptake, leaf water potential, shoot turgor pressure, osmotic potential, dry matter production, leaf area expansion, stomatal aperture size, stomatal diffusive conductance, and photosynthesis (Booker
et al., 1992; Chou and Lin, 1976; Einhellig et al., 1970; Einhellig and Kuan, 1971; Einhellig and Rasmussen, 1979; Einhellig et al., 1985; Gerald et al., 1992; Patterson, 1981). Allelochemicals have been reported to mediate the synthesis or degradation of some plant hormones, such as the activation of ABA synthesis by ferulic acid (Hollapa and Blum, 1991), the degradation of IAA via the stimulation of IAA oxidase by dihydroflavonone naringenin (Stenlid, 1970), and the resemblance of antiauxin and/or antigibberellin activity by sesquiterpene farnesol and sesquiterpene lactone argrophylline A and b_selinene (Komai et al., 1981; Watanabe et al., 1982).
Among so many symptoms, a decrease in photosynthesis efficiency is a common effect of allelopathic phenolics. Sorgoleone, a p-benzoquinone, in Sorghum bicolor root exudates was found to inhibit the oxygen evolution of soybean leaf disk and isolated pea chloroplast, which in turn caused growth reduction (Einhellig et al., 1993) and photosystem II electron transfer reaction (Gonzalez et al., 1997). The growth, chlorophyll content, and net photosynthesis of Lemna minor decrease as it is treated with juglone (Hejl et al., 1993). Benzoic and cin
*Corresponding author. Tel: 002-886-2-27821258 ext. 612; Fax: 002-886-2-27827954; E-mail: cmyang@gate.sinica.edu.tw